











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo

 Permalink
Permalink
Introducción
Puccinia triticina (Eriks), causante de la roya de la hoja (RH) del trigo (Triticum aestivum L.), es la enferme-dad más difundida del cultivo a nivel mundial1. La resistencia genética es la forma más económica y efectiva de controlarla. Los cultivares de trigo que son resistentes al momento de su liberación pueden tener niveles crecientes de infección debido a la aparición y selección de nuevas razas de este patógeno2.
Mains y Jackson3 fueron los primeros en demostrar la especialización fisiológica en P. triticina. Doce razas fueron distinguidas por los tipos de infección (TI) producidos en once diferenciales de trigo3. En los Estados Unidos de América (EUA) y Canadá, una se-rie de 12 líneas cercanamente isogénicas (NILS) derivadas de Thatcher (T. aestivum) se utilizan como diferenciales4. Estas líneas, cada una con un solo gen Lr (Lr, del inglés «Leaf rust») de resistencia, fueron desarrolladas por el Dr. Peter Dyck en el Centro de Investigación de Cereales, Agricultura y Agroalimentación de Canadá en Winnipeg, y se utilizan como diferenciales en la mayor parte del mundo5. Diversas instituciones de investigación han desarrollado sus propios siste-mas de análisis y designación de razas, pero algunas se unieron en el uso del sistema utilizado en Norteamérica, con el fin de facilitar la comunicación5.
Los estudios de la variabilidad del patógeno son importantes para determinar su rango de virulencia, para inferir el mecanismo de variación del patógeno, determinar el origen y dispersión de los patotipos, complementar el conocimiento de la resistencia de los cultivares comerciales, confirmar la presencia de nuevas razas inferidas por el cambio de comportamiento del hospedero e identificar razas importantes para el mejoramiento genético y estudios específicos6. La ventaja de contar con NILS con un solo gen de resistencia permite conocer directamente si el patógeno posee un gen complementario de avirulencia/virulencia a través de la reacción de resistencia/susceptibilidad del genotipo de trigo7. Para la designación de las razas se sigue la nomenclatura usada en Norte América y en América Latina descrita por Long y Kolmer4.
Se han conducido estudios de variabilidad de P. triticina en base a su fenotipo de virulencia por muchos años en los EUA, Canadá, Australia8, Brasil y Argentina9. Espe-cíficamente en Uruguay se han llevado estudios de identificación de razas desde el año 199110. El objetivo de este trabajo fue identificar las razas de Puccinia triticina pre- sentes en muestras de RH recolectadas en Uruguay durante 2011 y 2012.
Materiales y métodos
Aislamientos monopustulares de Puccinia triticina
Se estudiaron 205 aislamientos obtenidos de 158 muestras recolectadas durante el año 2011 y 145 aislamientos obtenidos de 117 muestras recolectadas durante el año 2012, provenientes de 97 cultivares y líneas diferentes de trigo y de ocho departamentos de la región triguera de Uruguay (Colonia, Soriano, Tacuarembó, Río Negro, Paysandú, Durazno, Florinda, Lavalleja).
Se sembraron 15 semillas del material susceptible Little Club en macetas de 10 cm de diámetro con una mezcla 1:1:1:1 de tierra, vermiculita, arena y sustrato (Biofer al- mácigos, Riverfilco; Biofer Ltd., Montevideo). Al producir- se la emergencia de las plántulas se aplicaron 20 mlde una solución de hidrácida maleica (Aldrich Chemical Co., Inc., WI, EUA) en la concentración de 0,36 g/l por maceta con el fin de reducir el desarrollo de las plantas y promover la producción de esporas. A los siete días, con la pri- mera hoja desarrollada, las plántulas fueron inoculadas con esporas provenientes de una muestra, utilizando una espátula2. Las plántulas se mantuvieron en cámara con 100 % de humedad y temperatura de 18 ºC aproximada- mente durante 14 a 16 h en condiciones de oscuridad. Posteriormente, las macetas se trasladaron al invernáculo con temperatura 20-25 ºC y seis a ocho horas de luz suplementaria (Son T 400w de sodio) a la luz natural. Una vez observados los puntos de infección, se cortaron las plántulas de cada maceta dejando dos hojas con una sola pústula en cada una, con el objetivo de evitar contaminación1. Dos semanas después de la inoculación se obtuvieron los aislamientos monopustulares, recolectando esporas de cada pústula individual en una cápsula de ge- latina utilizando un micro-recolector2.
Identificación de razas de Puccinia triticina
La identificación de las razas se realizó utilizando un set diferencial compuesto por 12 NILs derivadas de Thatcher (Tc), cada una con un gen de resistencia diferente: Lr 1, 2a, 2c, 3, 9, 16, 24, 26, 3ka, 11, 17, 304. Las líneas con Lr10, Lr20, Lr19 y Lr39 se utilizaron como diferenciales adicionales. A cada cápsula conteniendo las esporas de P. triticina se le agregó 0,8 ml de aceite mineral liviano Soltrol 170 (Phillips Petroleum Co., Borger, TX, EUA) y con la suspensión resultante se inocularon los sets diferenciales por aspersión cuando las plántulas tenían la primera hoja completamente expandida. La incubación y condiciones del invernáculo fueron similares a las descriptas para realizar los aislamientos monopostulares.
Doce días después de la inoculación, se evaluó el tipo de infección (TI) de acuerdo con la escala descripta por Stakman, Stewart y Loegering11 donde TI 0 = respuesta inmune, sin uredinios o necrosis; TI; (fleck) = lesiones necróticas sin esporulación; TI 1 = pequeños uredinios rodeados por necrosis; TI 2 = pequeños uredinios rodea- dos por clorosis; TI 3 = uredinios moderados sin clorosis o necrosis; TI 4 = uredinios grandes sin clorosis o necrosis2. Se utilizó + y - para indicar el tamaño más grande y más pequeño de los uredinios que el TI clásico, respectivamente1. La designación de las razas se realizó en base a la nomenclatura propuesta por Long y Kolmer4, seguida de -10, 20, 39 para indicar virulencia adicional sobre los genes Lr10, Lr20 y Lr39, respectivamente.
Frecuencia de razas de Puccinia triticina y de virulencia sobre los genes Lr
La frecuencia de cada raza se calculó por el número de aislamientos de cada raza en relación al número total de aislamientos de cada año. La frecuencia de aislamientos recolectados temprano durante el período del cultivo (previo 15 de noviembre), tarde (a partir del 16 de noviembre), al norte o sur del Río Negro también se refirió al número total de aislamientos recolectados cada año. La frecuencia de virulencia se calculó sumando la frecuencia de todas las razas virulentas sobre cada gen Lr.
Resultados
Se identificaron 17 razas presentes en el año 2011 y 18 en 2012 (Cuadro 1). El número de razas con frecuencia mayor a 10 % fue de dos y tres, mientras que el número de razas con frecuencia mayor a 5 % fue de cuatro y cinco, para los años 2011 y 2012, respectivamente.
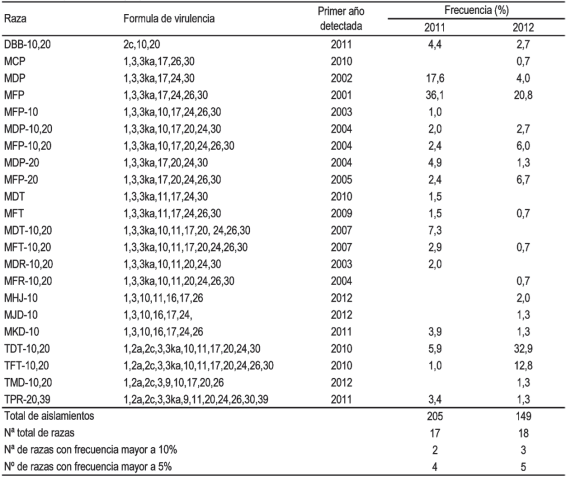
Cuadro 1: Raza, fórmula de virulencia, año de primera detección y frecuencia de las razas de Puccinia triticina identificadas durante 2011 y 2012 en Uruguay.
Todas las razas más frecuentes durante el período estudiado habían sido detectadas en años anteriores. La raza MFP fue la más frecuente durante 2011 y también fue una de las tres razas más frecuentes durante 2012. La raza MDP tuvo una frecuencia alta en el año 2011, sin embargo, en el año 2012 su frecuencia disminuyó a menos de 5 %. La raza TDT-10,20, tuvo una frecuencia relativamente baja en el año 2011, aumentando en importancia hasta llegar a ser la más frecuente durante el año 2012.
La raza TFT-10,20 presentó una tendencia similar, pasando de 1,0 % en 2011 a 12,8 % en 2012. MDT-10,20 tuvo una frecuencia intermedia durante 2011 y no fue detectada en 2012. Las razas MFP-10,20 y MFP-20 estuvieron presentes con una frecuencia de 2,4 % en 2011 e incrementaron su presencia en 2012 a niveles intermedios.
Otras razas identificadas antes del año 2011 estuvieron presentes generalmente en baja proporción (frecuencia menor a 5 %) en ambos años (MDP-10,20; MFT; MFT- 10,20), o en un año (MFP-10, MDT, MDR-10,20, MDT- 10,20 en 2011 y MCP, MFR-10,20 en 2012), excepto MDT-10,20 que tuvo una frecuencia de 7,2 % en el año 2011. También se identificaron seis razas que no habían sido detectadas previamente en Uruguay, DBB-10,20, MKD-10 y TPR-20,39 en el año 2011 y MHJ-10, MJD-10
y TMD-10,20 en el año 2012, todas en frecuencia menor a 5 %.
De acuerdo a la similitud de las razas estimada en base a su fórmula de avirulencia/virulencia (Figura 1), se observó un grupo de 16 razas similares que incluyen a MCP-, MDP-, MFP-, MDT-, MFT-, TDT-, TFT-. En este grupo, las razas con las letras D o F correspondiente al segundo sub-set son muy cercanas, ya que difieren en un solo gen de virulencia (Lr26). Otro grupo de tres razas está compuesto por MHJ-10, MJD-10 y MKD-10, las dos últimas se diferencian también por su reacción sobre el gen Lr26. Se detectaron tres razas más divergentes entre sí y con los grupos anteriores: TPR-20,39, TMD- 10,20 y DBB-10,20.
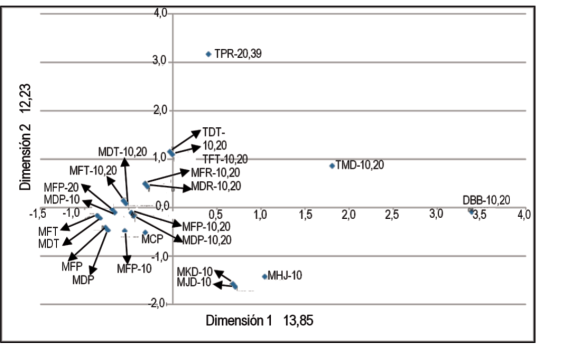
Figura 1: Análisis de correspondencia multivariada en base a la similitud de los genes de avirulencia/virulencia de las razas de Puccinia triticina.
Al analizar el momento de recolección de las muestras de las que se identificaron las razas más frecuentes (MDP, MFP y TDT-10,20), considerando sólo aquellas colectadas durante el período de cultivo, se observó una variación temporal importante. La raza MDP fue más frecuente en muestras recolectadas tardíamente (a partir del 16 de noviembre) en el año 2011 (Figura 2) y esta tendencia se invirtió en el año 2012. La raza MFP se detectó con alta frecuencia temprano en el 2011, no se detectó en muestras recolectadas tardíamente en ese año, mientras que en 2012 se detectó temprano con frecuencia intermedia, disminuyendo su frecuencia en muestras tardías. La raza TDT-10,20 se detectó temprano en el año 2011 y estuvo ausente en muestras recolectadas tardía- mente, sin embargo en el año 2012 apareció temprano con una frecuencia relativamente alta y su frecuencia disminuyó en muestras recogidas tarde.
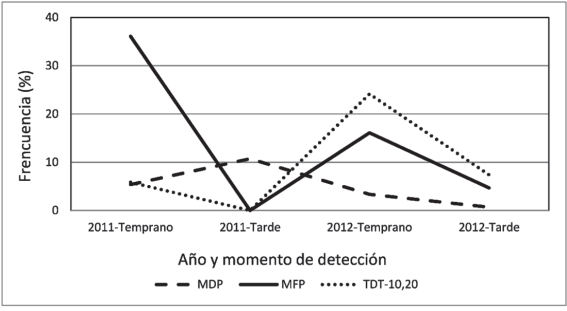
Figura 2: Frecuencia de las razas más frecuentes recolectadas en distintos momentos de 2011 y 2012. Temprano: hasta ell 15 de noviembre, tarde: a partir del 16 de noviembre.
Entre las razas poco frecuentes (frecuencia menor a 5 %), algunas sólo aparecieron en muestras recolecta- das temprano o tardíamente, como MDR-10,20 que sólo se identificó tardíamente en 2011 y MFP-10 sólo se identificó tempranamente en el mismo año (datos no mostrados). La misma situación se observó con las razas MFR-10,20 y MHJ-10, que sólo se encontraron en muestras tardías de 2012. La raza DBB-10,20, sólo se detectó temprano en el año 2011, mientras que en 2012 sólo se identificó tarde. MDP-10,20 se detectó temprano en 2011 y durante todo el período del cultivo en 2012. MDP-20 se observó en ambos años y momentos de recolección de muestras, pero en el 2011 la frecuencia fue de 3,9 % temprano, luego decreció, lo mismo ocurrió con la raza MKD-10. La raza TPR-20,39 apareció tarde en ambos años.
Considerando las razas con frecuencia mínima de 2,5 % en las zonas de recolección 2011 norte, 2011 sur, 2012 norte o 2012 sur, se observó que la detección de algunas razas dependió de su procedencia. Las razas MDP y MFP fueron identificadas en mayor frecuencia en muestras del año 2011 recolectadas en el sur, MDP-20 sólo fue detectada en el norte ambos años, y TDT-10,20 y TFT-10,20 fueron más frecuentes en muestras del año 2012 recolectadas en el norte (Figura 3). Otras razas no presentaron un patrón de aparición que pueda ser asocia- do a la zona de recolección.
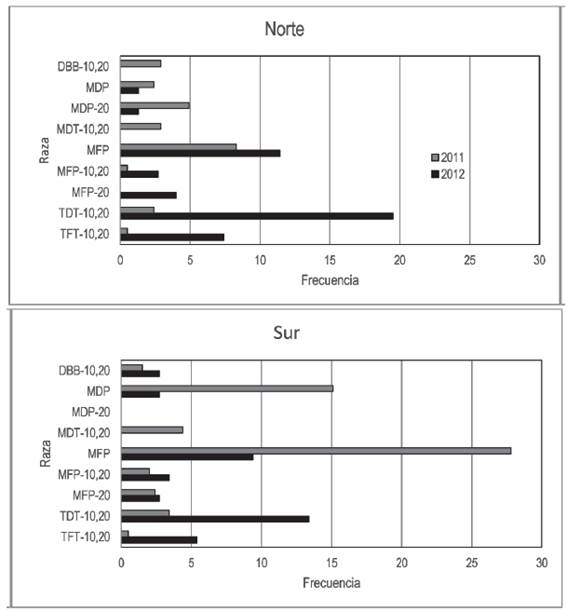
Figura 3: Frecuencia de razas de Puccinia triticina en el norte y sur de Uruguay durante 2011 y 2012 con frecuencia mínima de 2,5 % en al menos un período.
Se aislaron varias razas a partir de muestras provenientes de algunos materiales. Sin embargo, algunas fueron más específicas, como la raza DBB-10,20 que sólo se aisló del cultivar Génesis 2359 en los dos años, excepto por un aislamiento obtenido de Baguette 11 en 2011 (datos no presentados). El fenotipo MDR-10,20 fue aislado de INIATorcaza (tres aislamientos) y ORL 99192 (un aislamiento) durante 2011. MHJ-10 se identificó en una muestra del cultivar Buck Charrúa, una del material Thatcher y una de la NIL TcLr16 en 2012. La raza MJD-10 se aisló del cultivar Biointa 3000 durante el año 2012 (dos aislamientos). La raza MKD-10 fue detectada en muestras provenientes en su mayoría del cultivar Biointa 3004. TMD-10,20, se identificó en una muestra de TcLr9, mientras que TPR-20,39 se identificó principalmente de mues- tras de la línea con Lr39 en ambos años.
Todos los aislamientos obtenidos de INIA Carpintero correspondieron a la raza MFP, excepto un aislamiento de 2012 caracterizado como MDP. Similarmente, la mayoría de los aislamientos provenientes de Baguette 11 y LE 2381-Génesis 2381 fueron MFP (2011 y 2012), como cuatro aislamientos obtenidos a partir de Klein Pantera y tres de Baguette 17 (2011). La única muestra analizada del cultivar IPR 84 resultó ser la raza MFP-10-20. Las razas aisladas de las NILS TcLr2a y TcLr2c de 2012 fue- ron TDT-10,20 y TFT-10,20, respectivamente.
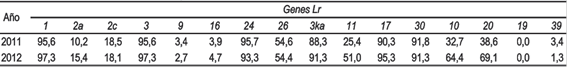
La frecuencia de virulencia sobre los genes Lr1, Lr3, Lr24, Lr3ka, Lr17 y Lr30 fue alta, sobre los genes Lr2a, Lr2c, Lr26, Lr11, Lr10 y Lr20 fue intermedia, sobre los genes Lr9, Lr16 y Lr39 fue baja y no se detectó ninguna raza virulenta sobre Lr19 (Cuadro 2).
Discusión
Los resultados obtenidos en este trabajo indican cambios importantes en la población de P. triticina entre los años estudiados. Esto se evidenció tanto en la frecuencia de las razas ya reportadas como en la aparición de las razas no reportadas previamente DBB-10,20, MHJ-10, MJD-10, MKD-10, TMD-10,20, TPR-20,39, lo que indica
una gran variación del patógeno, coincidiendo con lo señalado por otros investigadores1)(2)(12. No obstante, mu- chas de las razas identificadas fueron reportadas hace más de 10 años y aún persisten en los cultivos de trigo.
Junto a TDT-10,20, las razas más frecuentes fueron MFP y MDP. Estas últimas son similares en su fórmula de avirulencia/virulencia, difiriendo sólo en su comporta- miento sobre el gen Lr26 que condiciona una reacción intermedia, evaluada como TI 2 o 2+ en el caso de ailsamientos avirulentos o un TI 3 o superior para aislamientos considerados virulentos. Esta variación podría también ser causada por distintas condiciones ambienta- les dentro del invernáculo, principalmente temperatura. La sensibilidad a la temperatura también fue reportada para los genes Lr11, Lr16, Lr17, Lr18 y Lr2313. La poca diferenciación por la reacción frente a Lr26 también ocurre con otros pares de razas muy próximas en el análisis de correspondencia. En Argentina, las razas MDP y MFP fueron frecuentes durante el año 201014 y 201115, lo que confirma la similaridad de las poblaciones de P. triticina entre Uruguay y Argentina16. Muchos aislamientos identificados como MFP fueron obtenidos del cultivar Baguette 11, uno de los cultivares más sembrados durante los años 2011-201217. Esto podría explicar su amplia prevalencia, ya que, en general, la frecuencia relativa de cada raza responde al área de siembra de los cultivares susceptibles9. MFP también es virulenta sobre otros cultivares difundidos y fue la raza presente en todas las muestras de INIA Carpintero recolectadas en el año 2011. Este cultivar fue resistente hasta 2009, cuando ocupaba un área de siembra moderada, y presentó altos niveles de RH en 2010. La raza asociada a este cambio de comportamiento fue una variante de la raza MFP que adquirió virulencia sobre este cultivar y constituyó la mayoría de los aislamientos de esta raza a partir de su primera detección en el año 2009. MFP probablemente adquirió virulencia sobre un gen de resistencia presente en INIA Carpintero que no está incluido en el set diferencial utilizado para este estudio.
La raza MDP se identificó en los dos momentos de recolección de muestras considerados, siendo detectada más frecuentemente muestras tardías del año 2011, probablemente por multiplicación de su inóculo en los sucesivos ciclos de infección. La raza MFP fue detectada en tres momentos de muestreo y no se identificó en muestras recolectadas tardíamente en el año 2011. Es probable que los cultivares sembrados tardíamente en 2011 fueran resistentes a esta raza o el área foliar de los cultivares susceptibles se haya desecado prematuramente (antes del 15 de noviembre). MDP y MFP fueron más frecuentes en aislamientos de muestras recolectadas en el sur en el año 2011, lo que podría deberse a una mayor concentración de cultivares susceptibles a estas razas en esa zona.
Las otras razas cercanas integrantes del mismo grupo MDP/MFP con virulencia sobre Lr10 y/o Lr20 surgieron en los años 2004 y 200512. Probablemente estas deriva- ron de MDP y MFP, por mutación de avirulencia a virulencia sobre los genes mencionados. MFP-10, identifica- da por primera vez en el año 2004, sólo se obtuvo a partir de muestras de la línea NT103 del año 2011.
MDP-20 y MFP-20 se detectaron en muestras de distintos cultivares de trigo en ambos años de este estudio. Estas razas fueron inicialmente identificadas en el año 200418, asociadas a altas infecciones de RH en el cultivar INIA Tero, previamente resistente, y han prevalecido en las chacras desde su primera detección. La raza MDP- 20 se detectó sólo en el norte mientras que MFP-20 se presentó en el sur sólo en 2011, lo que podría deberse a limitantes de muestreo, ya que se trata de razas relativa- mente poco frecuentes.
MDP-10,20 y MFP-10,20, detectadas en ambos años de estudio en diferentes cultivares, también han aparecido todos los años desde su primera detección. MDT sólo se detectó en aislamientos de muestras colectadas durante el año 2011 y MFT se detectó en 2011 y muestras colectadas en el sur en 2012. MDT y MFT podrían haber surgido de las razas MDP y MFP por mutación de avirulencia a virulencia sobre Lr11.
MDT-10,20 y MFT-10,20 se identificaron por primera vez en el año 200718. Ambas estuvieron presentes todos los años durante 2007-2010. En el presente trabajo, la primera se detectó a partir de muestras colectadas de distintos cultivares sólo en el año 2011. En Brasil, estas razas se identificaron por primera vez en el año 200419 y desde el norte posiblemente hayan migrado a Uruguay12.
TDT-10,20 y TFT-10,20 fueron identificadas por primera vez en el año 201018. Estas razas combinan el mayor número de genes de virulencia entre las razas encontradas. La frecuencia de ambas incrementó en 2012, y fue- ron aisladas de varios cultivares comerciales como Arex, Atlax, Baguette 13, Biointa 1002, Biointa 3000, Biointa 3006, Buck Charrúa e INIA Tero. Son virulentas también sobre otros cultivares altamente difundidos como Baguette 11 y Baguette 19, lo que explicaría su prevalencia. Probablemente provienen de Brasil16 donde fueron inicialmente identificadas en 200519, y han prevalecido desde su primera detección. TDT-10,20 no se registró en muestras tardías del año 2011, probablemente por las mismas razones discutidas para la raza MFP.
MDR-10,20, identificada inicialmente en el año 2003, estuvo presente en la población de P. triticina desde su detección hasta el año 2008. Esta raza fue responsable del cambio de comportamiento de los cultivares INIA Churrinche e INIATorcaza20. MFR-10,20, detectada inicialmente en el año 2004, estuvo presente sólo en el año 2012.
La raza DBB-10,20, detectada por primera vez en Uruguay en el período comprendido en este estudio, es similar en términos de virulencia a BBB-10,20 detectada en Argentina sobre trigos duros en el año 2001 y 2002 (P. Campos, comunicación personal, 30 de mayo de 2012). Es posible entonces que DBB-10,20 haya surgido de BBB-10,20 adquiriendo virulencia sobre el gen de resistencia Lr2c (única diferencia). Esta raza afectó a Génesis 2359, que presentó muy buen comportamiento frente a RH hasta el año 2010, siendo muy resistente en el estado de plántula a todas las razas probadas hasta ese momento18. Durante 2010 se detectaron infecciones de severidad intermedia en chacras en la zona noreste, y más tardíamente en algunas chacras del litoral del país, pero en forma muy heterogénea. El año 2011 fue un año atípico dado que la infección de RH comenzó tarde y en general tuvo un desarrollo menor al de otros años, principalmente al sur del área de siembra, probablemente por la falta de humedad libre sobre el follaje necesaria para la infección del patógeno.
Las nuevas razas MJD-10, MHJ-10 y TMD-10,20 difieren de aquellas reportadas hasta 2012 en tres o más genes de avirulencia/virulencia, por lo que no es posible identificar un precursor directo en Uruguay desde 198912)(21, ya sea proveniente de Argentina en base a los datos del período 2002-201214 o de Brasil en el período 2002-200919. Las razas MKD-10, MJD-10 y MJH-10
son similares y se caracterizan por ser las únicas con virulencia sobre el gen Lr16, que es frecuente en cultiva- res argentinos14. MKD-10 y MJD-10 difieren sólo por su reacción sobre el gen Lr26. La raza TMD-10,20 es virulenta sobre Lr9 y Lr24.
TPR-20,39, virulenta sobre Lr9 y Lr39, se encontró en muestras de TcLr9 y TcLr39, lo que corresponde con su virulencia específica sobre estos genes. La virulencia sobre Lr9 y Lr39 también se presentó en Argentina con una frecuencia de 2 %, y fue reportada por primera vez en el año 201014.
Las mismas razas se reproducen y dispersan dentro de una zona epidemiológica. Uruguay está comprendido en la zona que también abarca a las regiones trigueras de Argentina, Brasil y Paraguay22. Dentro de esta región, en general, las poblaciones del P. triticina presentes en Argentina y Uruguay son muy similares23, probablemente debido a la similitud de los cultivares sembrados en ambos países. La misma situación de cultivares similares y probable presencia de razas similares de P. triticina ocurre entre Brasil y Paraguay18. Las razas que afectan a una amplia gama de cultivares se propagan muy rápido dentro de toda la zona epidemiológica. También puede existir dispersión de razas a nivel intercontinental, como se confirmó para la raza MCD-10,20 que probablemente migró desde México hacia América del Sur23. Se podría inferir que la alta frecuencia de virulencia sobre Lr1, Lr3, Lr24, Lr3ka, Lr17 y Lr30 pueda estar asociada a la presencia de estos genes en los cultivares más sembrados en Uruguay y en la región.
Este trabajo confirma la aparición continua de razas nuevas de P. triticina en Uruguay, la mayoría de las cuales probablemente evolucionaron a través de mutaciones o migración y posterior selección por su virulencia sobre los cultivares sembrados16.

