










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Links relacionados
Compartilhar

 Permalink
PermalinkAgrociencia (Uruguay)
versão impressa ISSN 1510-0839versão On-line ISSN 2301-1548
Agrociencia Uruguay vol.20 no.2 Montevideo dez. 2016
Suplementación con glicerina cruda y afrechillo de arroz entero a vacas de carne pastoreando campo natural
Clariget Juan Manuel1, Pérez-Clariget Raquel2, Álvarez-Oxiley Andrea2, Bentancur Oscar3, Bruni María de los Ángeles3
1Instituto Nacional de Investigación Agropecuaria (INIA) La Estanzuela. Ruta 50, km 11, Colonia, Uruguay.
Correo electrónico: jclariget@inia.org.uy.
2Universidad de la República, Facultad de Agronomía. Avenida Garzón 780, 12900 Montevideo, Uruguay.
3Universidad de la República, Facultad de Agronomía. Ruta 3 km 363, 60000 Paysandú, Uruguay.
Recibido: 21/8/15 Aceptado: 14/9/16
Resumen
Con el objetivo de evaluar los efectos de la suplementación con glicerina cruda (GC) y afrechillo de arroz entero (AA) sobre la dinámica ruminal, la cinética de degradación de la materia seca (MS), el consumo de forraje y variables plasmáticas asociadas al balance energético en vacas de carne pastoreando campo natural, cuatro vacas fistuladas en el rumen fueron asignadas a un cuadrado latino 4 × 4 con un período adicional. Los tratamientos evaluados fueron: suplementación con GC (550 mL/vaca/día), suplementación con AA (1kg de MS/vaca/día), suplementación con GC + AA (550 mL/vaca/día + 1 kg de MS/vaca/día) y sin suplementación (CON). La asignación diaria de forraje para todos los tratamientos fue 10 % del peso vivo. La GC asociada o no al AA disminuyó el pH ruminal en las primeras seis horas pos-suplementación, pero no afectó ningún parámetro de degradabilidad de la MS del forraje. Comparada con CON la suplementación con GC disminuyó el consumo de forraje pero no afectó el consumo de MS total ni de energía metabolizable (EM). Ni el consumo de forraje, ni el consumo total fueron diferentes entre la suplementación con GC + AA y AA, sin embargo el consumo de EM fue mayor para GC + AA en comparación con el resto. La suplementación con GC + AA aumentó la concentración plasmática de glucosa e insulina sin afectar la de B-hydroxibutirato. De los suplementos ensayados, la mezcla GC + AA (mayor nivel de suplementación) logró los mejores resultados biológicos.
Palabras clave: RUMEN, METABOLISMO, GLICEROL, ENERGÍA
Summary
Supplementation of Beef Cows Grazing Natural Pasture with Crude Glycerin and Whole Rice Bran
To study the effects of supplementation with crude glycerin (GC) and whole rice bran (AA) on ruminal dynamics, kinetics of dry matter (DM) degradation, forage intake and plasma variables associated with energy balance in beef cows grazing natural pasture, four rumen fistulated cows were allocated to a latin square 4 × 4 with an additional period. The treatments were: supplementation with GC (550 mL/cow/day), supplementation with AA (1 kg DM/cow/day), supplementation with GC + AA (550 mL/cow/day + 1 kg DM/cow/day) and without supplementation (CON). The daily forage allowance for all treatments was 10 % of live weight. GC associated or not to AA decreased ruminal pH on the first six hours pos-supplementation, but did not affect any parameters of degradability of forage DM. GC supplementation decreased forage intake but did not affect total DM intake and metabolizable energy (ME) compared with CON. GC + AA and AA supplementation did not differ on intake of forage and total DM, however the ME intake was higher for GC + AA compared to the rest. GC + AA supplementation increased plasma concentration of glucose and insulin without affecting B-hydroxibutirate. Among the supplements tested, GC + AA mixture (higher level of supplementation) achieved the best biological results.
Keywords: RUMEN, METABOLISM, GLYCEROL, ENERGY
Introducción
La producción de biodiesel deja disponible para la alimentación animal un importante volumen de glicerina cruda (GC). La misma se produce en el orden del 10 % del biodiesel elaborado (Larosa, 2001). La GC contiene glicerol, agua, lípidos, cenizas y metanol (Schröder y Südekum, 1999), cuya proporción depende del proceso industrial. El glicerol, componente principal de la GC, es reconocido como un ingrediente seguro para la alimentación animal por la legislación de EEUU (GPO, 2016) y de Europa (Alexander et al., 2010).
Una vez suministrado, el glicerol llega al rumen donde tiene tres posibles destinos: la fermentación, la absorción o continuar sin ser atacado por los micro-organismos (Krehbiel, 2008). El glicerol incrementa la producción de ácidos grasos volátiles (AGV) tanto in vivo (Rémond et al., 1993; Mach et al., 2009) como in vitro (Trabue et al., 2007; Ferraro et al., 2009; Bruni et al., 2013); fundamentalmente aumenta la producción de ácidos propiónico y butírico (Ferraro et al., 2009; Wang et al., 2009a). Cuando la producción de ácido butírico se incrementa se observa un aumento de la concentración plasmática de B-hydroxibutirato (BHB; Rémond et al., 1993). Tanto el propionato producido por la fermentación del glicerol como el glicerol absorbido directamente como tal por las paredes del rumen, tienen como destino el hígado, donde son sustratos de la neoglucogénesis. El control de la gluconeogénesis tiene un componente endocrino en el que se destaca la hormona insulina responsable de la homeostasis de la glucosa. Esta hormona promueve la captación celular de glucosa y su oxidación y puede disminuir la gluconeogénesis hepática en rumiantes (Brockman y Laarveld, 1986).
La mayor parte de la literatura internacional está referida al uso del glicerol en vacas lecheras (Chung et al., 2007; Wang et al., 2009b; Carvalho et al., 2011). También se ha utilizado como integrante de la dieta en el engorde de ganado bovino (Mach et al., 2009; Parsons et al., 2009) y de ovinos (Gunn et al., 2010), en terneros (Gunn et al., 2011) y en vaquillonas de re-emplazo (Moriel et al., 2010). Sin embargo, son escasos los reportes sobre su utilización en ganado de cría en pastoreo. Existen reportes sobre aumentos del consumo (Bodarski et al., 2005), disminución (De Frain et al., 2004) o ausencia de cambios observables (Donkin et al., 2009) cuando se agregó GC a la dieta de vacas lecheras. Por otra parte, no se han observado cambios en la digestibilidad de la materia seca (MS) y de la fibra detergente neutra (FDN) cuando el glicerol se incorporó a la dieta de ovinos (Schröder y Südekum, 1999) o bovinos (Donkin et al., 2009). Sin embargo, Wang et al. (2009b) han planteado que la administración de glicerol podría aumentar la tasa de degradabilidad de la MS y todos los parámetros de degradabilidad de la FDN, mientras que Shin et al. (2012) sugieren que la digestibilidad de la primera no se vería modificada pero la digestibilidad de la fibra se vería disminuida. De Frain et al. (2004) reportan disminuciones del pH ruminal cuando incorporan glicerol a la dieta de vacas lecheras sin encontrar cambios en la concentración de nitrógeno amoniacal (N-NH3). La disminución del pH ruminal podría explicar al menos parcialmente la menor digestibilidad de la fibra (Calsamiglia y Ferret, 2002) reportada por Shin et al. (2012).
El afrechillo de arroz entero (AA), subproducto de uno de los principales cultivos de la región, es considerado un alimento energético con buen aporte de proteínas y minerales (White y Hembry, 1985; Wang et al., 2012). El mismo tiene un alto contenido de grasa (15 %; NRC, 2000), por lo que alimentación en exceso a rumiantes disminuye la digestión de la fibra (Palmquist y Jenkins, 1980). Niveles de suplementación del 1 % Peso Vivo (PV), disminuyen el consumo de forraje sin afectar la digestibilidad del mismo ni el consumo total de MS (Gadberry et al., 2004). Su inclusión sobre pasturas o henos de calidad media a niveles de 50 % de la dieta total disminuyen el pH ruminal e incrementan la concentración de AGV y de N-NH3 (Cárdenas-García et al., 1992). El mismo presenta una buena palatabilidad para los animales y está disponible en el mercado regional. Constituye así una alternativa para utilizar en suplementaciones en rodeos de cría (Pérez-Clariget et al., 2007; Quintans et al., 2012; Astessiano et al., 2013; Soca et al., 2013) ya sea solo o mezclado con glicerina cruda dado que esta última es líquida a temperatura ambiente.
La hipótesis planteada es que la GC administrada a vacas adultas pastoreando campo natural disminuiría el pH ruminal, incrementando la concentración plasmática de glucosa, insulina y BHB, sin afectar el consumo de forraje ni la degradabilidad de la MS del forraje tanto cuando se la administra sola como cuando se la agrega al AA. El objetivo del presente trabajo fue evaluar los efectos de la suplementación con GC asociada o no al AA, sobre la dinámica ruminal estimada a través del pH y la concentración de N-NH3, la cinética de degradación de la MS, el consumo de forraje y la concentración plasmática de glucosa, insulina y BHB en vacas de carne pastoreando campo natural.
Materiales y métodos
El protocolo experimental fue aprobado por la Comisión de Ética en el Uso de Animales (CEUA). El experimento se llevó a cabo en la Estación Experimental «Bernardo Rosengurtt» (EEBR), Facultad de Agronomía, Universidad de la República (Udelar) Cerro Largo (latitud 32Ú21’.20 S, longitud 54Ú26’.32 O), entre mayo y setiembre 2013.
Animales, tratamiento y diseño experimental
Se utilizaron cuatro vacas Angus y Angus x Hereford, vacías, fistuladas en el rumen, con más de seis unidades de condición corporal (CC; escala: 1-8 unidades; Vizcarra et al., 1986) y 604 ± 43 kg PV, las cuales se asignaron a un cuadrado latino 4 × 4 con un período adicional (cinco períodos en total: P1–P5). Los cuatro tratamientos evaluados fueron: suplementación con glicerina cruda (550 mL/vaca/día; GC); suplementación con afrechillo de arroz entero (1kg de MS/vaca/día; AA); suplementación con glicerina cruda (550 mL/vaca/día) + afrechillo de arroz entero (1 kg MS/vaca/día; GC+AA) y sin suplementación (CON). Cada periodo de suplementación abarcó 18 días: 12 días de adaptación a la dieta (días 1 al 12) y seis días de muestreos (días 13 al 18).
Pasturas, asignación de forraje y suplementos
Durante todo el experimento las vacas pastorearon en conjunto sobre campo natural con una asignación de forraje de 10 % PV. La disponibilidad del forraje fue determinada por el método de doble muestreo (Haydock y Shaw, 1975) a través de un cuadrado de 50 cm x 50 cm, con cinco puntos en la escala y dos repeticiones, cortando el forraje al ras del suelo. La altura del mismo se registró como la altura de la última hoja que tocara la regla. Por apreciación visual del cuadrado de muestreo se estimó la relación verde/seco (Rel V/S; Cuadro 1). Los animales se pesaban al inicio de cada período y en base a la disponibilidad y el peso de los animales se determinaba el área a pastorear para ese período.

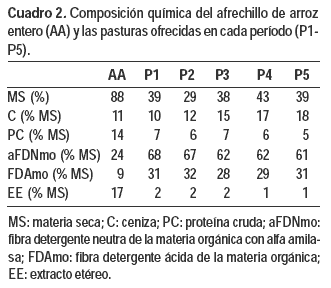
La composición química del afrechillo de arroz y de las pasturas ofrecidas por período (Cuadro 2) fueron evaluadas a través de análisis químico [% MS, % extracto etéreo (AOAC, 1990; N.167.03, N.954.02, respectivamente), % cenizas, % proteína cruda (AOAC, 2007; N.942.05, N.984.13; respectivamente), % fibra detergente neutra (con alfa amilasa) y ácida de la materia orgánica con tecnología ANKOM de forma secuencial (Van Soest et al., 1991)]. La composición de la glicerina cruda fue: 3 % agua (AOCS, 2009; Ea8-58), 6 % cenizas (AOCS, 1973; Ea2-38), 77 % glicerol (AOCS, 2012; Ea6-51), 13 % materia grasa (AOAC, 1980; N.14.019) y su contenido de metanol fue aportado por ALUR (Alcoholes del Uruguay; 1 %). La energía metabolizable (EM) aportada por la glicerina cruda y por el afrechillo de arroz entero fueron tomadas de los cuadros FEDNA (FEDNA, 2010), la energía de las pasturas fue estimada a partir de la digestibilidad in vitro de la MS (DMS) utilizando la ecuación (EM (Mcal/kg MS) = (4,4 x 0,82 x % DMS)/ 100) de Mahanna (1990).
Muestreos y determinaciones
Se determinó la degradabilidad in situ de la pastura y el consumo de forraje durante los días 13, 14 y 15 de cada periodo experimental. Para estimar la degradabilidad in situ de la MS de la pastura se utilizó la técnica de Mehrez y Ørskov (1977). Se colocaron 20 bolsas ANKOM (20 cm × 10 cm; tamaño promedio de poros: 50 micrones) por vaca en cada período experimental en las que se colocaron 4 g de MS de una muestra representativa de forraje utilizando la técnica de hand-clipping (Weir y Torell, 1959). El forraje fue previamente picado para lograr un tamaño de 5 mm. Las bolsas se introdujeron en el saco ventral del rumen todas juntas a la hora 0 (7:00 am) y se extrajeron por duplicado en 10 horarios (0, 2, 4, 8, 12, 16, 24, 36, 48 y 72 h) de incubación en el rumen. Cada bolsa fue lavada bajo corriente de agua fría hasta obtener un líquido incoloro. Luego se secaron en estufa a 60 ºC durante 48 h y se pesaron para obtener el residuo no degradado. Para cada vaca, período y tratamiento se utilizó el siguiente modelo (Ørskov y McDonald, 1979):
Di = a + b (1 – e-c t )
donde:
Di: Degradabilidad al tiempo ti (%)
a: Fracción soluble (%)
b: Fracción potencialmente degradable (%)
c: Tasa de degradación (%/h)
t: Tiempo en horas
Con los parámetros de degradabilidad (a, b y c) y asumiendo una tasa de pasaje (Kp) de 2 %/h y 4 %/h se estimó la degradabilidad efectiva (DE) aplicando la siguiente ecuación:
DE = a + [bc / (c+Kp)].
El consumo de materia seca del forraje (CMSf) se determinó con la técnica de doble marcador (Penning, 2004). Para esto se empleó óxido de cromo (Cr2O3) como marcador externo y la fibra detergente ácido indigestible (FDAi) como indicador interno. Dos veces por día se dosificaron 7,5 g de Cr2O3 por un periodo de 10 días: siete días para estabilizar el marcador y tres días para recolección de heces. A partir del día 7 se recolectaron muestras de heces directamente desde el recto dos veces por día, las que fueron posteriormente secadas en estufa de aire forzado a 60 °C hasta peso constante y molidas en molino con malla 1 mm. Luego por animal y periodo de muestreo se determinó la concentración de Cr por espectrofotometría de absorción atómica (Silva y Queiroz, 2006). Se tomaron muestras del forraje ofrecido (hand-clipping) en cada periodo de determinación del consumo. Las muestras fueron secadas en estufa de aire forzado a 60 ºC hasta peso constante. Las muestras de forraje, suplementos y heces recolectadas fueron molidas con malla de 2 mm y se sometieron a una prueba de digestibilidad in situ durante 144 h. Una vez finalizado el procedimiento de incubación in situ, se retiraron los residuos correspondientes a cada muestra y se conservaron hasta la realización del análisis de la FDAi (Van Soest et al., 1991). Finalmente para estimar el CMSf se utilizó la siguiente ecuación:
CMSf (kg/vaca/d) = ([FDAih]*H/0,8 – [FDAis]*CMSs)/ [FDAif]
donde:
CMSf: Consumo de materia seca del forraje
FDAih: Porcentaje de FDAi en las heces
H: Heces excretadas
0,8: Recuperación de la FDAi en las heces, según Sunvold y Cochran (1991)
FDAis: Porcentaje de FDAi en el suplemento
CMSs: Consumo de materia seca del suplemento
FDAif: Porcentaje de FDAi en el forraje
A las: 0, 0:30, 1:30, 3, 6, 12 y 24 h de comenzada la suplementación (07:00 am) de los días 17 y 18 se tomaron muestras de sangre de la vena yugular utilizando tubos heparinizados para la determinación de la concentración plasmática de glucosa, insulina y BHB. Luego de extraídas, las muestras fueron centrifugadas a 1529 g durante 15 minutos y el plasma almacenado a -20 ºC. La insulina se determinó por análisis inmunoradiomético (IRMA) en fase sólida (Diasource, Bruselas, Bélgica). Los coeficientes de variación intraensayo para el control bajo (23 uUI/mL) y alto (86,5 uUI/mL) fueron 7,5 y 6,9 %, respectivamente y el límite de detección fue de 0,52 uUI/mL. Las concentraciones de BHB y glucosa se determinaron por espectrofotometría utilizando kits comerciales (D-3 Hydroxibutyrate, Randox, Reino Unido y Glucose-Glucose oxidasa/Peroxidasa, ByoSistem S.A, Barcelona, España) con coeficientes de variación intraensayo de 6,5 y 7,7 %, respectivamente.
Los días 17 y 18 también se realizaron extracciones de líquido ruminal, a las: 0, 0:30, 1:30, 3, 6, 9, 12, 16 y 24 h de comenzada la suplementación (07:00 am). Las muestras extraídas fueron filtradas en tela doble de lienzo (quesería) y se determinó el pH inmediatamente después de extraídas. Posteriormente se tomaron muestras para determinar N-NH3, que fueron congeladas a -20 ºC hasta su posterior análisis. La determinación de N-NH3 se realizó por medio de destilación directa con Kjeldahl (Galyean, 2010).
Análisis estadístico
Las concentraciones de pH, N-NH3, glucosa, BHB e insulina se analizaron con un modelo de medidas repetidas en el tiempo que incluyó como efecto aleatorio el animal y como efecto fijo el período, tratamiento, hora e interacción tratamiento por hora. El consumo de materia seca del forraje, materia seca total y energía metabolizable se analizaron con el mismo modelo pero sin el efecto fijo de la hora y la interacción tratamiento por hora. Los parámetros de degradabilidad de la MS se analizaron con el modelo no lineal (PROC NLIN) propuesto por Ørskov y MCDonald (1979). Los parámetros se compararon por intervalos de confianza. Los resultados se expresaran en medias ajustadas ± error estándar (ee) y las diferencias estadísticamente significativas se consideraran con P ≤ 0,05 y tendencia con P ≤ 0,10.
Modelo:
Yijkl= b0 + Vi + Pj + Tk + R1 + R2 + R3 + R4 + b1Xijk + eijk + Hl + (TH)kl + eijkl
donde:
b0: intercepto
Vi: efecto animal
Pj: efecto periodo
Tk: efecto tratamiento
R1: efecto residual del tratamiento 1
R2: efecto residual del tratamiento 2
R3: efecto residual del tratamiento 3
R4: efecto residual del tratamiento 4
b1: coeficiente de regresion de la covariable Xijk (valor inicial de la variable medida)
eijk: error experimental
Hl: efecto hora de medición
(TH)kl: interacción tratamiento por hora
eijkl: error de la medida repetida
Resultados
Consumo y parámetros de degradabilidad del forraje
La suplementación con GC, AA y GC + AA disminuyó (P < 0,01) 21 %, 14 % y 7 % el consumo de forraje en comparación con el tratamiento CON, respectivamente (Cuadro 3), pero sólo el valor obtenido con GC fue estadísticamente diferente (P < 0,05) al observado en CON. El consumo de MS total de GC y AA fue 15 % y 5 % menor que CON, mientras que la suplementación con GC + AA fue 8 % mayor; sin embargo ninguna de estas diferencias fueron significativas (Cuadro 3). No se encontraron diferencias en la cantidad de energía metabolizable ingerida entre CON y GC o AA. El mayor (P<0,05) consumo de energía metabolizable se obtuvo en GC + AA (Cuadro 3).
Los parámetros de degradabilidad de la MS del forraje (a: fracción soluble, b: fracción potencialmente degradable, c: tasa de degradación) no fueron afectados por el tipo de suplemento (P > 0,10). La DE tanto al 2 %/h como al 4 %/h tampoco difirió entre tratamientos (P > 0,10; Cuadro 3).
pH y N-NH3
El tratamiento afectó el pH ruminal (P = 0,03); el promedio diario fue mayor (P < 0,05) en CON que en AA, mientras que los valores obtenidos cuando se suplementó con GC o GC + AA fueron intermedios (Cuadro 4). Se encontró una interacción tratamiento por hora (P = 0,04); en efecto, cuando los animales fueron suplementados el pH del rumen fue inferior (P < 0,05) comparado con CON a las horas: 1,5, 3 y 6, sin que se encontraran diferencias entre las distintas suplementaciones (Figura 1a). Los valores de pH en todos los casos fueron mayores a 6,6.
El N-NH3 no fue afectado por los tratamientos (P = 0,30; Cuadro 4). Se encontró una interacción tratamiento por hora (P = 0,04); el N-NH3 de los animales suplementados con AA y GC + AA fue mayor (P < 0,05) a la hora: 0,5 de terminada la suplementación comparados con el tratamiento CON; mientras que los suplementados con GC presentaron concentraciones intermedias. A partir de la hora 6 y hasta las 16, el tratamiento CON presentó los mayores (P < 0,05) valores de N-NH3, sin que se encontraran diferencias entre los tratamientos suplementados (Figura 1b).
B-hydroxibutirato, glucosa e insulina
La concentración de BHB no fue afectada por el tratamiento (P = 0,78) ni por la hora (P = 0,69) y tampoco se encontró interacción tratamiento por hora (P = 0,40). El tratamiento afectó la concentración plasmática diaria de glucosa (P = 0,02) e insulina (P < 0,01). La suplementación con GC+AA aumentó 5 % (P < 0,05) la concentración plasmática diaria de glucosa y 24 % (P < 0,01) la de insulina comparado con el tratamiento CON, mientras que las suplementaciones con GC o AA presentaron concentraciones intermedias (Cuadro 4). La hora no influyó (P = 0,28) en la concentración plasmática de glucosa, pero se encontró una tendencia (P = 0,09) a que fuera afectada por la interacción tratamiento por hora (Figura 2). A la hora 0,5 la concentración plasmática de glucosa fue mayor (P < 0,05) cuando se suplementó con GC + AA en comparación con el tratamiento CON, mientras que la suplementación con GC y AA presentaron concentraciones intermedias (Figura 2a).
La concentración plasmática de insulina fue influenciada por la hora (P < 0,01) y por la interacción tratamiento por hora (P < 0,01). A las horas 0,5 y 1,5, la concentración plasmática de insulina fue mayor (P < 0,05) en GC + AA en comparación con CON, mientras que la suplementación con GC y AA presentó concentraciones intermedias (Figura 2b).
Discusión
La suplementación con GC + AA aumentó el consumo de EM y la concentración plasmática de glucosa e insulina, logrando los mejores resultados biológicos. Sin embargo, hay que tener en cuenta que los suplementos evaluados en el presente trabajo no fueron iso-energéticos, siendo GC + AA el de mayor nivel. De todos modos, el aporte de los tres suplementos fue bajo en relación al PV de los animales (0,1-0,3 % PV), aunque similares (levemente inferiores) a varios trabajos nacionales evaluando al AA como suplemento en vacas de cría (0,4-0,6 % PV; Pérez-Clariget et al., 2007; Quintans et al., 2012; Astessiano et al., 2013; Soca et al., 2013). Se ha reportado que la utilización de carbohidratos rápidamente fermentecibles a niveles similares a los utilizados en este trabajo (menos de 10 % en la dieta total) como suplementos de forrajes de mediana o baja calidad produce efectos asociativos positivos ente el forraje y el suplemento. Esta acción estaría mediada por la estimulación de la actividad microbiana que impactaría positivamente en la digestión del forraje y la síntesis de proteína a nivel del rumen, lo que se conoce como «efecto starter» (Opatpatanakit et al., 1993; Dixon y Stockdale, 1999). El efecto starter de los suplementos utilizados en el presente trabajo no es claro ya que no se observó un aumento de la degradabilidad del forraje con ninguno de los tres suplementos utilizados.
La disminución en el consumo de forraje observada cuando se suplementó solo con GC es coincidente con los resultados obtenidos por De Frain et al. (2004) trabajando con vacas de leche durante el período seco. Sin embargo, no coincide con el trabajo de Almeida et al. (2013) realizado en vacas primíparas sobre pasturas tropicales. Moore et al. (1999) en una revisión de 66 trabajos concluyó que suplementaciones energéticas cuando el forraje base tiene una relación de Nutrientes Digestibles Totales (NDT) / PC mayor a 7 disminuye el consumo de forraje. Para este trabajo se estimó una relación de 7,8 (47,5 / 6,1) utilizando la composición químicas de las pasturas y calculando los NDT en base a NRC (2000). Posiblemente esta sea una de las razones por las cuales la suplementación energética únicamente (GC) haya disminuido el consumo, dado que al compararla con el trabajo de Almeida et al. (2013) presentaba una relación de 4,5 (60 / 13,3), por lo cual la suplementación energética no afectó el consumo de forraje. Por otra parte, se ha sugerido que la disminución del consumo producida por la suplementación con glicerol estaría asociada al poder neoglucogénico del mismo (De Frain et al., 2004). Es posible que el aumento de la concentración de ácido propiónico que produce la fermentación ruminal del glicerol (Rémond et al., 1993) actuando en el propio rumen, así como el posterior incremento de la concentración de insulina, puedan ser responsables de la disminución del consumo voluntario (Allen, 2000). Bodine y Purvis (2003) pastoreando con novillos forrajes de 6 - 8 % PC y suplementando solamente con concentrado energético, o con concentrado energético + proteico, observaron que la suplementación únicamente con concentrado energético disminuyó el consumo de forraje pero cuando la suplementación fue con ambos concentrados el consumo de forraje fue similar al presentando por el grupo control. Estos resultados concuerdan con los obtenidos en el presente trabajo donde la suplementación con AA o AA + GC no disminuyó el consumo de forraje, posiblemente debido al agregado de PC en la dieta.
La mayoría de los trabajos realizados en vacas lecheras en lactación (Donkin et al., 2009; Wang et al., 2009a) o ganado de engorde (Fisher et al., 1973; Mach et al., 2009) no observan cambios en el consumo total (ración totalmente mezclada; RTM) cuando se sustituyeron concentrados energéticos (maíz o cebada) por glicerol o cuando lo incorporaron directamente. Sin embargo, Bodarski et al. (2005) trabajando con vacas lecheras observaron aumentos del consumo total cuando agregaron glicerol a la RTM. Ellos lo atribuyen al exceso de proteína en la dieta de las vacas, dado que el agregado de glicerol en la RTM mejoró la fermentación ruminal incrementándose la síntesis de proteína microbiana y por ende el consumo total. En este trabajo no se observaron diferencias en el consumo total al agregar glicerol en la dieta cuando se compara CON vs, GC o AA vs, AA + GC, resultados similares a los obtenidos por la mayoría de los trabajos publicados.
Los resultados obtenidos en el presente trabajo avalan la hipótesis que el glicerol actuaría sobre el consumo voluntario a través de señales metabólicas asociadas al propiónico e insulina sin modificar los parámetros de degradabilidad de la MS. En efecto, al igual que lo reportado por otros autores (Schröder y Südekum, 1999; Hess et al., 2008; Donkin et al., 2009; Almeida et al., 2013) no se observaron cambios en la degradabilidad del forraje con ninguno de los suplementos utilizados. Si bien se observó una disminución del pH ruminal en las primeras seis horas pos-suplementación cuando se suministraron todos los suplementos evaluados, los valores registrados se encontraban dentro del rango 6,6 a 7,1. Ørskov (1982), Mould et al. (1983) y Calsamiglia y Ferret (2002), indican que un pH ruminal debajo de 6,2 reduce la actividad de bacterias celulolíticas y la digestión de forrajes groseros, indicando que depresiones en el pH podrían ser responsables de las reducciones en la digestibilidad de la fibra asociada con la suplementación energética. En el presente trabajo los valores de pH estuvieron siempre por encima de 6,6 por lo cual era esperable no afectar los parámetros de degradabilidad del forraje del campo natural por la inclusión de los suplementos. Otros autores (Rémond et al., 1993; De Frain et al., 2004) también reportaron una disminución del pH ruminal cuando incorporaron glicerol a la dieta dentro de los mismos rangos de valores registrados en el presente trabajo.
En coincidencia con lo publicado por De Frain et al. (2004) no se observaron cambios en la concentración ruminal de N-NH3 cuando se agregó glicerol a la dieta. La ausencia de cambios en N-NH3 está en concordancia con el bajo nivel de PC que presenta la glicerina cruda. Los valores medios registrados de N-NH3 en todos los tratamientos (entre 9-11 mg/100 mL) se encontraban dentro del rango (8-30 mg/100 mL) considerado óptimo para el funcionamiento ruminal. Valores inferiores a 5 mg/100 mL se han asociado a dietas con déficit en proteína o con resistencia a la degradación de la misma, mientras que niveles superiores a 30 mg/100 mL serían causados por dietas con excesos de proteína o déficit de energía (McDonald et al., 1986).
Se han reportado rápidos incrementos de la glucemia después de la administración de glicerol en vacas lecheras. En efecto, Goff y Horst (2001) observaron incrementos de 16 a 25 % en los primeros 30 minutos, luego de la administración de 1 a 3 L de glicerol como tratamiento para prevenir la cetosis en vacas lecheras. Wang et al. (2009a) trabajando con vacas lactando encontraron aumentos de glucosa de 9 % administrando 300 g de glicerol a la dieta en comparación con las control (sin glicerol), realizando los sangrados dos horas después de brindada la comida. Ambos autores coinciden con lo reportado en el presente trabajo, donde se encontraron aumentos en la concentración plasmática de glucosa en la primera media hora luego de finalizada la suplementación. Este rápido incremento de la concentración de la glucosa estaría asociado a la rápida desaparición en el rumen de los suplementos utilizados. Más del 80 % del glicerol desaparece en las primeras dos (Kijora et al., 1998) o cuatro horas (Rémond et al., 1993) pos-suplementación. Mach et al. (2009) trabajando con toros en terminación e incluyendo hasta 8 % de glicerol en la dieta encontraron incrementos de 45 % en la concentración plasmática de insulina, resultados similares o algo superiores a los de este trabajo. Por lo cual, debido a que aumenta la concentración de glucosa o porque aumenta la concentración de insulina, lo que ambas variables plasmáticas reflejan es una mejora en el estatus energético provocado por la suplementación.
Wang et al. (2009a) encontraron en vacas lactando disminuciones de BHB suplementando con 300 g de glicerol en comparación con las control. La diferencia encontrada en BHB, Wang et al. (2009a) la plantean como positiva ya que lo que buscaban era disminuir la cetosis, la cual se ve reflejada con menores concentraciones de BHB. En el presente trabajo, el hecho de no encontrase diferencias en la concentración de BHB parece lógico ya que el mismo podría haber aumentado por efecto de la dieta (dietas con glicerol pueden incrementar la concentración de ácido butírico (Shin et al., 2012) y el mismo se puede ver reflejado en aumentos de BHB plasmático (Rémond et al., 1993), o podría haber disminuido dado que el suplemento podría haber cubierto las demandas energéticas y de esta forma presentar un mejor balance energético (Wang et al., 2009a). Por lo cual el no verse afectada la concentración de BHB parece razonable.
Conclusiones
La suplementación con glicerina cruda no afectó los parámetros de degradabilidad de la MS del forraje, la concentración N-NH3 ruminal y de BHB plasmática. La suplementación con GC + AA presentó los mayores consumos de energía y las mayores concentraciones plasmáticas de glucosa e insulina. Estos resultados muestran que a niveles de 0,1 % PV es posible la utilización de glicerina cruda como suplemento de vacas de carne pastoreando campo natural, sin mostrar ningún perjuicio aparente. Su administración junto con el AA es una alternativa viable a nivel de campo y bajo esta forma (GC + AA) fue donde se obtuvieron los mejores resultados biológicos.
Agradecimientos
A la Ing. Agr. PhD Graciela Quintans por prestarnos las vacas con las cuales se realizó este trabajo. A la ANII por la beca de maestría (POS_2011_1_3522) y proyecto (FMV_3_2011_1_6713) otorgada para realizar y financiar parte de este trabajo. A ALUR por brindarnos la glicerina cruda, a Cousa por realizarnos los análisis químicos de la glicerina cruda y a todo el personal de la EEBR, en particular a Baltasar Martínez y al Ing. Agr. Carlos Mantero.
Bibliografía
Alexander J, Benford D, Boobis A, Ceccatelli S, Cravedi JP, Doerge D, Domenico A, Dogliotti E, Edler L, Farmer P, Filipic M, Fink-Gremmels J, Fürst P, Guerin TK, Knutsen H, Machala M, Mutti A, Schlatter J, Van-Leeuwen R. 2010. Scientific opinion on the abiotic risks for public and animal health of glycerine as co-product from the biodiesel production from category 1 animal by-products (ABP) and vegetable oils. European Food Safety Authority, 8(12): 1 – 22.
Allen M. 2000. Effects of Diet on Short-Term Regulation of Feed Intake by Lactating Dairy Cattle. Journal of Dairy Science, 83(7): 1598 – 1624.
Almeida H, Pereira M, Olivera E, Bacelar F, Abreu G, Andrade A, Texeira P, Santos P. 2013. Crude glycerin in supplement to primiparous lactating cows grazing on tropical pasture: nutritional and productive characteristics. Revista Brasileira de Zootecnía, 42(2): 117 – 124.
AOAC. 2007. Official Methods of Analysis. 18th ed. Washington : Association of Official Analytical Chemists. 1v. (en varias paginaciones).
AOAC. 1990. Methods of Analysis. 15th ed. Arlington : Association of Official Analytical Chemists. 2v.
AOAC. 1980. Methods of Analysis. 13th ed. Washington : Association of Official Analytical Chemists. 1018p.
AOCS. 2012. 103rd AOCS Annual Meeting and Short Courses [En línea]; April 29 - May 2, 2012; Long Beach, California, USA. Consultado 12 octubre 2016. Disponible en: https://www.aocs.org/attend-meetings/archives/103rd-aocs-annual-meeting-and-short-courses.
AOCS. 2009. 100th AOCS Annual Meeting and Short Courses [En línea]. May 3-6, 2009; Rosen Shingle Creek, Orlando, Florida, USA. Consultado 12 octubre 2016. Disponible en: https://www.aocs.org/attend-meetings/archives/100th-aocs-annual-meeting-and-short-courses.
AOCS. 1973. Teachnical program AOCS 64th annual spring meeting New Orleans, Louisiana April 29-May 2, 1973. Journal of the American Oil Chemists’ Society, 50(2): A73-A79.
Astessiano AL, Pérez-Clariget R, Espasandín AC, López-Mazz C, Soca P, Carriquiry M. 2013. Metabolic, productive and reproductive responses to postpartum short-term supplementation in primiparous beef cows. Revista Brasileira de Zootecnia, 42(4): 246 – 253.
Bodarski R, Werteleki T, Bommer F, Gosiewski S. 2005. The changes of metabolic status and lactation performance in dairy cows under feeding RTM with glycerine (glycerol) supplement at periparturient period. Electronic Journal of Polish Agricultural Universities, 8(4): 22.
Bodine TN, Purvis HT. 2003. Effects of supplemental energy and/or degradable intake protein on performance, grazing behavior, intake, digestibility, and fecal and blood indices by beef steers grazed on dormant native tallgrass prairie. Journal Animal Science, 81: 304 – 317.
Brockman RP, Laarveld B. 1986. Hormonal regulation of metabolism in ruminants : a review. Livestock Science, 14: 313 – 334.
Bruni M, Galindo J, Aldama A, Moreira O, Chilibroste P. 2013. Efecto del nivel de glicerol sobre de fermentación ruminal in vitro de ensilaje de pradera [Cd-Rom]. En: XXIII Reunión de ALPA y IV Congreso Internacional de Producción Animal Tropical, La habana, Cuba.
Calsamiglia S, Ferret A. 2002. Fisiología ruminal relacionada con la patología digestiva : acidosis y meteorismo [En línea]. En: XVIII Curso de especializacion FEDNA. pp. 97-115.Consultado 22 setiembre 2016. Disponible en: http://www.produccion-animal.com.ar/sanidad_intoxicaciones_metabolicos/metabolicas/metabolicas_bovinos/27-acidosis_meteorismo.pdf.
Cárdenas-García D, Newbold CJ, Galbraith H, Topps JH. 1992. The effect of including colombian rice polishings in the diet on rumen fermentation in vitro. Animal Production, 54(2): 275 – 280.
Carvalho ER, Schmelz-Roberts NS, White HM, Doane PH, Donkin SS. 2011. Replacing corn with glycerol in diets for transition dairy cows. Journal Dairy Science, 94: 908 – 916.
Chung YH, Rico DE, Martinez CM, Cassidy TW, Noirot V, Ames A, Varga GA. 2007. Effects of feeding dry glycerine to early postpartum Holstein dairy cows on performance and metabolic profiles. Journal Dairy Science, 90: 5682 – 5691.
De Frain JM, Hippen AR, Kalscheur KF, Jardon PW. 2004. Feeding glycerol to transition dairy cows : effect on blood metabolites and lactation performance. Journal Dairy Science, 87: 4195 – 4206.
Dixon RM, Stockdale CR. 1999. Associative effects between forages and grains : consequences for feed utilisation. Australian Journal Agricultural Research, 50(5): 757 – 773.
Donkin, SS, Koser SL, White HM, Doane PH, Cecava MJ. 2009. Feeding value of glycerol as a replacement for corn grain in rations fed to lactating dairy cows. Journal Dairy Science, 92: 5111 – 5119.
FEDNA. 2010. Tablas FEDNA de composición y valor nutritivo de alimentos para la fabricación de piensos compuestos [En línea]. 3a ed. Madrid : FEDNA. 502p. Consultado 6 octubre 2016. Disponible en: http://www.fundacionfedna.org/tablas-fedna-composicion-alimentos-valor-nutritivo.
Ferraro SM, Mendoza GD, Miranda LA, Gutiérrez CG. 2009. In vitro gas production and ruminal fermentation of glycerol, propylene glycol and molasses. Animal Feed Science and Technology, 154: 112 – 118.
Fisher LJ, Erfle JD, Lodge GA, Sauer FD. 1973. Effects of propylene glycol or glycerol supplementation of the diet of adiry cows on feed intake, milk diet and composition, and incidence of ketosis. Canadian Journal of Animal Science, 53: 289 – 296.
Gadberry MS, Beck PA, Gunter SA. 2004. Forage intake and performance by beef heifers grazing cool-season pasture supplemented with de-oiled rice bran or corn. The Professional Animal Scientist, 20(5): 394 – 400.
Galyean ML. 2010. Laboratory Procedures in Animal Nutrition Research [En línea]. Lubbock : Texas Tech University. Consultado 12 octubre 2016. Disponible en: https://www.depts.ttu.edu/afs/home/mgalyean/lab_man.pdf.
Goff JP, Horst RL. 2001. Oral glycerol as an aid in the treatment of ketosis/fatty liver complex. Journal Dairy Science, 84(1): 153.
GPO. 2016. §582.1320 : Glycerin [En línea]. En: Electronic Code of Federal Regulations [En línea] Consultado 6 octubre 2016. Disponible en: http://www.ecfr.gov/cgi-bin/text-idx?SID=cab8db5852cdb66a57b44e75b982ca87&mc=true&node=se 21.6.582_11320&rgn=div8.
Gunn PJ, Lemenager RP, Claeys MC, Lake SL. 2011. Effects of distiller’s dried grains with soluble and crude glycerin on performance and carcass characteristics in early weaned beef calves. The Professional Animal Scientist, 27: 283 – 294.
Gunn PJ, Schultz AF, Van Emon ML, Neary MK, Lemenager RP, Rusk CP, Lake SL. 2010. Effects of elevated crude glycerin concentrations on feedlot performance, carcass characteristics, and serum metabolite and hormone concentrations in finishing ewe and wether lambs. The Professional Animal Scientist, 26: 298 – 306.
Haydock KP, Shaw NH. 1975. The comparative yield method for estimating dry matter yield of pasture. Australian Journal of Experimental Agriculture and Animal Husbandry, 15: 663 – 670.
Hess B W, Lake SL, Gunter SA. 2008. Using glycerin as a supplement for forage-fed ruminant. Journal Animal Science, 86: 392.
Kijora C, Bergner H, Gotz KP, Bartelt J, Szakacs J, Sommer A. 1998. Research note : investigation on the metabolism of glycerol in the rumen of bulls. Archivos Tierernaehrung, 51(4): 341 – 348.
Krehbiel CR. 2008. Ruminal and physiological metabolism of glycerin. Journal Animal Science, 86: 392.
Larosa RJ. 2001. Proceso para la producción de BIODIESEL(metilester o esteres metílicos de ácidos grasos) : Refinación de glicerina [En línea]. 8p. Consultado 20 abril 2010. Disponible en: http://www.biodiesel-uruguay.com/articulos/Biod-rev2.pdf.
Mach N, Bach A, Devant M. 2009. Effects of crude glycerine supplementation on performance and meat quality of Holstein bulls fed high-concentrate diets. Journal Animal Science, 87: 632 – 638.
Mahanna WC. 1990. Pioneer Forage Manual : A Nutritional Guide. Iowa : Pioneer Hi-Bred International. 55p.
McDonald P, Edwrds RA, Greenhalg JF. 1986. Digestión. En: Nutrición Animal. Zaragoza : Acribia. pp. 131 – 156.
Mehrez AZ, Ørskov ER. 1977. A study of the artificial fiber bag technique for determining the digestibility of feeds in the rumen. Journal Agricultural Science, 88: 645 – 650.
Moore JE, Brant MH, Kunkle WE, Hopkins DI. 1999. Effects of supplementation on voluntary forage intake, diet digestibility, and animal performance. Journal Animal Science, 77(2): 122 - 135.
Moriel P, Nayigihugu V, Cappellozza BI, Gonçalves JM, Krall T, Foulke KM, Cammack KM, Hess BW. 2010. Camelina meal and crude glycerin as feed supplements for developin replacement beef heifers. Journal Animal Science, 89: 4314 – 4324.
Mould FL, Ørskov ER, Mann SO. 1983. Associative effects of mixed feeds : I. Effects of type and level of supplementation and the influence of the rumen fluid pH on cellulolisis in vivo and dry matter digestion of various roughages. Animal Feed Science and Technology, 10: 15 – 30.
NRC. 2000. Nutrient requirement of beef cattle. 7a ed. Washington : NRC. 232p.
Opatpatanakit Y, Kellaway RC, Lean IJ. 1993. Substitution effects of feeding rolled barley grain to grazing dairy cows. Animal Feed Sciencie and Technology, 42: 25 – 38.
Ørskov ER. 1982. Host animal protein requirement and protein utilization. En: Protein Nutrition in Ruminants. London : Academic Press. pp. 85–135.
Ørskov ER, McDonald I. 1979. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. The Journal of Agricultural Science, 92: 499 – 503.
Palmquist DL, Jenkins TC. 1980. Fat in lactation rations : Review. Journal of Dairy Science, 63: 1 – 14.
Parsons GL, Shelor MK, Drouillard JS. 2009. Glycerin improves performance of feedlot heifers when fed at low levels [En línea]. En: Plains Nutrition Council Spring Conference, San Antonio, USA. Consultado 12 de octubre. Disponible en: http://amarillo.tamu.edu/files/2010/11/2008PNC-Proceedings1.pdf.
Penning PD. 2004. Herbage intake handbook. 2da ed. Reading : The British Grassland Society. 191p.
Pérez-Clariget R, Carriquiry M, Soca P. 2007. Estrategias de manejo nutricional para mejorar la reproducción en ganado bovino. Archivos Latinoamericanos de Producción Animal, 15: 114 – 119.
Quintans G, Scarsi A, Banchero G. 2012. Short-term supplementation with rice bran in pre-partum primiparous grazing beef cows. En: 63° Congreso Europeo de Producción Animal; 27 - 31 agosto, 2012; Bratislava, Eslovaquia. Wageningen : Wageningen Academic Publishers. pp 77.
Rémond B, Souday E, Jouany JP. 1993. In vitro and in vivo fermentation of glycerol by rumen microbes. Animal Feed Science and Technology, 41: 121 – 132.
Schröder A, Südekum KH. 1999. Glycerol as a by-product of biodiesel production in diets for ruminants [En línea]. Consultado 22 setiembre 2016. Disponible en: http://regional.org.au/au/gcirc/1/241.htm.
Shin JH, Wang D, Kim SC, Adesogan AT, Staples CR. 2012. Effects of feeding crude glycerin on performance and ruminal kinetics of lactating Holstein cows fed corn silage or cottonseed hull-based, low-fiber diets. Journal Dairy Science, 95: 4006 – 4016.
Silva J, Queiroz AC. 2006. Análise de alimentos : métodos químicos e biológicos. 3ra ed. Vicosa : UFV. 235p.
Soca P, Carriquiry M, Keisler DH, Claramunt M, Do Carmo M, Olivera-Muzante J, Rodríguez M, Meikle A. 2013. Reproductive and productive response to suckling restriction and dietary flushing in primiparous grazing beef cows. Animal Production Science, 53: 283 – 291.
Sunvold GD, Cochran RC. 1991. Technical note : evaluation of acid detergent lignin, alkaline peroxide lignin, acid insoluble ash, and indigestible acid detergent fiber as internal markers for prediction of alfalfa, bromegrass, and prairie hay digestibility by beef steers. Journal Animal Science, 69: 4951 – 4955.
Trabue S, Scoggin K, Tjandrakusuma S, Rasmussen MA, Reilly PJ. 2007. Ruminal fermentation of Propylene Glycol and Glycerol. Journal of Agricultural Food Chemestry, 55: 7043 – 7051.
Van Soest PJ, Robertson JB, Lewis BA. 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. Journal Dairy Science, 74(10): 3583 – 3597.
Vizcarra JA, Ibañez W, Orcasberro R. 1986. Repetibilidad y reproductividad de dos escalas para estimar la condición corporal de vacas Hereford. Investigaciones Agronómicas, 7: 45 - 47.
Wang C, Liu Q, Yang WZ, Huo WJ, Dong KH, Huang YX, Yang XM, He DC. 2009a. Effects of glycerol on lactation performance, energy balance and metabolites in early lactation Holstein Dairy cows. Animal Feed Science and Technology, 151: 12 – 20.
Wang C, Liu Q, Yang WZ, Huo WJ, Yang WZ, Dong KH, Huang YX, Guo G. 2009b. Effects of glycerol on rumen fermentation, urinary excretion of purine derivatives and feed digestibility in steers. Livestock Science, 121: 15 – 20.
Wang Y, Xin HS, Li YZ, Zhang WW, Xia K, Wang ZB, Li M, Zhang YG. 2012. The effects of different processing methods on the estimated nutritional value of rice bran according to NRC-2001 Model or DVE/OEB System. Journal of Animal Feed Science, 21: 503 – 520.
Weir WC, Torell DT. 1959. Selective grazing by sheep as shown by a comparison of the chemlcal composition of range and pasture forage obtained by hand clipping and that collected by esophageal-fistulated sheep. Journal Animal Science, 18: 641 – 649.
White TW, Hembry FG. 1985. Rice by-products in ruminant rations. [Baton Rouge, La.] : Louisiana Agricultural Experimental Statation. 18p.

