










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Links relacionados
Compartilhar

 Permalink
PermalinkAgrociencia (Uruguay)
versão impressa ISSN 1510-0839versão On-line ISSN 2301-1548
Agrociencia Uruguay vol.18 no.2 Montevideo dez. 2014
Caracterización y descripción de los estadios ninfales de Borellia bruneri (Rhen, 1906) (Orthoptera: Gomphocerinae)
Miguel Lucía1, Lorier Estrellita1, Zerbino Stella2
1Facultad de Ciencias, Universidad de la República (Udelar). Iguá 4225, 11400 Montevideo, Uruguay
2INIA (Instituto Nacional de Investigación Agropecuaria) La Estanzuela, Colonia 70000, Uruguay
Correo electrónico: lorier@fcien.edu.uy
Recibido: 25/9/13 Aceptado: 21/8/14
Resumen
En la primavera del año 2008 en la región centro-sur del Uruguay se registró un importante incremento en la población de Borellia bruneri que no se registraba desde hacía 50 años. El momento adecuado de realizar tratamientos para evitar daños es cuando la mayor proporción de la población se encuentra en el estadio de ninfa tres, así la eclosión de huevos es máxima, y se evita el momento de mayor consumo y de disminución en la sensibilidad a los insecticidas. Este trabajo fue realizado en 2009-2010 para conocer el ciclo y la fenología de esta especie en un pastizal natural en el departamento de Florida, Uruguay (33°45’9.30"S 55°54’11.90"O). Desde principios de octubre hasta mediados de enero se realizaron muestreos semanales, y luego muestreos quincenales hasta el fin del período a fines de abril. Los estadios ninfales se determinaron según el desarrollo de los rudimentos alares, la genitalia externa y el número de segmentos antenales. Complementariamente se realizó un estudio morfométrico para caracterizar los estadios previamente diferenciados por las características morfológicas. Se determinaron cinco estadios ninfales fácilmente distinguibles en base al desarrollo de los rudimentos alares y la genitalia externa. Largo de la cabeza, diámetro mayor del ojo y largo del fémur posterior fueron los caracteres más útiles para separar los individuos de los distintos estadios del mismo sexo y entre sexos. Esta información tiene utilidad para planificar estrategias de control oportunas sin que ocurran daños irreparables durante explosiones poblacionales de esta especie.
Palabras clave: BORELLIA BRUNERI, ESTADIOS NINFALES, CAMPO NATURAL
Summary
Characterization and Description of Borrellia bruneri (Rhen, 1906) (Orthoptera: Gomphocerinae) Nymphal Instars
In the spring of 2008 in the central-south region of Uruguay, there was a significant increase in the population density of Borrellia bruneri not witnessed for 50 years. The proper time for treatments to prevent damage is when the highest proportion of the population is in nymph III, so hatching eggs is maximum, and avoiding the peak of consumption and the decrease in sensitivity to insecticides. This work was conducted in 2009-2010 and aimed to identify the cycle and phenology of this species in a natural grassland located in Florida, Uruguay (33° 45' 9.30" S 55°54’11.90" O). From early October to mid January we conducted weekly sampling, and then biweekly until the end of the period in late April. The nymphal stages were determined according to the development of wing rudiments, external genitalia and the number of antennal segments. Additionally a morphometric study was performed to characterize the instars previously differentiated by morphological characters. Five easily distinguishable nymphal instars were determined, based on the development of the wing pads and external genitalia. Head length, the larger diameter of the eye, and length of the hind femur were the most useful characters to separate individuals of different nymphal instars of the same sex and between sexes. This information is useful for planning timely control strategies without irreparable damage during population explosions of this kind.
Keywords: BORELLIA BRUNERI, NYMPHAL INSTARS, NATURAL GRASSLAND
Introducción
En la primavera del año 2008, principalmente en los departamentos de la región centro-sur del Uruguay, se registró un importante incremento de la densidad poblacional de tucuras, particularmente de Borellia bruneri (Rehn, 1906). La composición de las comunidades en esta oportunidad fue similar a la de las explosiones poblacionales reportadas en el siglo pasado en la década de los 50´ (Carbonell, 1957; Lorier et al., 2010). En ambos momentos B. bruneri fue la especie más abundante, con densidades poblacionales que superaron varias veces a las de las otras especies presentes (Liebermann y Piran, 1941; Liebermann y Ruffinelli, 1946; Ruffinelli y Carbonell, 1953; Carbonell, 1957; Silveira Guido et al., 1958).
Esta especie se distribuye en todo el territorio nacional (Silveira Guido et al., 1958; COPR, 1982; Carbonell, 1995; Carbonell et al., 2006). Tiene preferencia por ambientes secos, posiciones topográficas más altas y vegetación baja, asociada a comunidades vegetales donde prevalecen las gramíneas de porte bajo. Estudios en distintas comunidades vegetales de la región Pampeana demuestran una alta asociación de B. bruneri con vegetación del tipo halófilo (de Wysiecki et al., 2004; Torrusio et al., 2002; Mariottini et al. 2012, 2013).
La dinámica de las poblaciones de las tucuras es altamente variable, no sigue direcciones simples, las respuestas no son lineales y resultan de efectos combinados de las fuerzas abióticas (clima), de las interacciones bióticas (competencia, depredación, parasitismo), del tipo y estructura de la vegetación, de los disturbios producidos por el hombre (pastoreo, fertilización, agricultura, forestación, quema sobre la pradera natural, uso de insecticidas) y de las características propias de la especie (ciclos supra-anuales) (Cigliano et al., 1995; Joern, 2000). Bajo ciertas condiciones favorables del clima, suelo y vegetación suelen hacer explosiones demográficas. Se han identificado relaciones entre factores abióticos y los distintos estados del ciclo de vida de los acridios, en general los inviernos relativamente secos y veranos también secos y con altas temperaturas favorecen la aparición de estos fenómenos demográficos. En los inviernos con déficit hídrico hay sobrevivencia alta de huevos, mientras que las lluvias frecuentes aumentan la mortalidad, principalmente por ataque de entomopatógenos. Las temperaturas altas aceleran el desarrollo. Con respecto a B. bruneri, de Wysiecki et al. (2011) estiman que las temperaturas cálidas y la vegetación rala afectan positivamente su demografía y su densidad.
Esta especie fue reportada como una de las más destructivas de las praderas, por los daños importantes que causó en el campo natural y las praderas artificiales (COPR, 1982; Carbonell, 1995; 1957; Lorier y Zerbino, 2009; Lorier et al., 2010). Históricamente fue una especie poco importante en Argentina (Carbonell, 1995; de Wysiecki et al., 2000), a partir de los seguimientos realizados en el año 2004 se le consideró una especie de distribución intermedia (de Wysiecki et al., 2004), y en los seguimientos de los últimos años (2005-2010) resultó ser una especie de amplia distribución y una de las más abundantes en la Provincia de Buenos Aires (Mariottini et al. 2012, 2013).
Estos insectos tienen metamorfosis gradual, completan su ciclo a través de tres estados: huevo, ninfa y adulto. Los huevos son depositados en el suelo, generalmente, en paquetes, reunidos por una sustancia semental esponjosa. En este estado tiene un período de detenimiento del desarrollo que se asocia con el período invernal o sequías estacionales (COPR, 1982; Hewitt y Onsager, 1983; Pfadt, 2002). Cuando en primavera la temperatura y humedad del suelo alcanzan el valor umbral, los embriones continúan su desarrollo, nacen las ninfas, que son semejantes a los adultos en su aspecto general, pero que se distinguen fácilmente de estos por el menor tamaño del cuerpo y por el grado de diferenciación y posición de las tecas alares. También se diferencian por el menor desarrollo de la genitalia externa y por el número de segmentos antenales (Zolessi 1956, 1958; Santoro y Carames, 1973; Santoro, 1976; Santoro et al. 1978; Turk y Barrera, 1976; Pfadt, 2002; Resh y Cardé, 2003; Bentancourt et al., 2009). Antes de alcanzar el estadio adulto, las ninfas mudan varias veces y la cantidad de estadios depende de factores bióticos o abióticos. En las especies de cinco estadios ninfales, a los primeros tres se les denomina «mosquitas» y a las ninfas mayores (IV y V) «saltonas». A partir del estadio IV se produce la inversión alar (Santoro et al., 1978).
En nuestro país se conoce poco acerca de la fenología de esta especie. Las primeras ninfas son observadas en octubre y si bien se reporta que el período ninfal tiene una duración de 40 a 50 días, existen diferencias entre autores en el número de estadios ninfales que tiene esta especie. Silveira Guido et al. (1958) mencionan que tiene seis estadios, mientras que recientemente Mariottini (2009) en Argentina determinó cinco estadios ninfales. Los adultos copulan en enero y febrero, y luego en los meses de marzo y abril las hembras comienzan a depositar sus huevos a una profundidad de (3,5-4) cm en suelos sin remoción y compactos, frecuentemente asociados a las raíces de Cynodon dactylon (COPR, 1982). Algunos autores consideran que es una especie univoltina (COPR, 1982; Mariottini et al., 2011), mientras que Martínez (comunicación personal) plantea que en Uruguay tendría dos generaciones por año.
Para el seguimiento sistemático de las poblaciones de tucuras, a fin que los técnicos puedan tomar decisiones de manejo adecuadas y planificar las estrategias de control oportunas, sin que ocurran daños irreparables, es necesario conocer el ciclo y la fenología de una especie (Berry et al. 1996; Sánchez y de Wysiecki, 1993). Con el objetivo de aportar información sobre el momento adecuado de realizar un control químico que resulte efectivo sobre las tucuras, en este trabajo se determina el número de estadios ninfales de B. bruneri y se describen las variables morfológicas y morfométricas a partir de individuos colectados en el campo.
Materiales y métodos
Durante la primavera-verano 2009-2010, entre principios de octubre y fines de abril, se realizó el estudio de la fenología de B. bruneri. El sitio de estudio seleccionado fue una pradera natural localizada en el departamento de Florida (33°45’9.30"S 55°54’11.90"O). La frecuencia de los muestreos fue semanal hasta mediados de enero y posteriormente quincenal hasta el final del periodo, totalizando 14 recolectas.
La segada con una red entomológica (aro de 42 cm de diámetro, red de 70 cm profundidad y mango de 120 cm) fue el método de muestreo utilizado. Se realizaron 200 golpes en una transecta al azar, de aproximadamente 3 m de ancho y 200 m de largo. Cada golpe de red consistió en un arco de 180º. El material colectado fue trasladado al laboratorio en bolsas plásticas dónde se lo congeló y posteriormente se fijó en alcohol al 70 %.
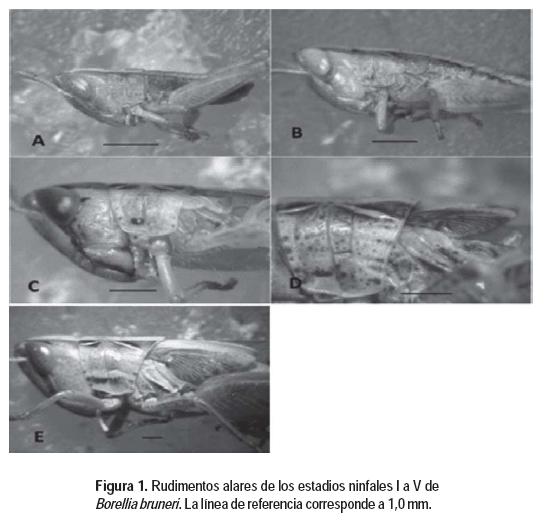
La identificación de la especie se realizó utilizando la bibliografía disponible (Silveira Guido et al., 1958; Carbonell, 1995; Carbonell et al., 2006) y a partir de la comparación con los ejemplares depositados en la Colección de Entomología de la Facultad de Ciencias de la Udelar. La clasificación de los estadios ninfales de B. bruneri se realizó mediante la comparación con los adultos bajo un microscopio estereoscópico Nikon ZM10 y utilizando la información de la bibliografía sobre el desarrollo postembrionario de otras especies de acridios (Zolessi, 1958; Turk y Barrera, 1976, 1979; Santoro et al., 1978). La obtención de las fotografías de ninfas enteras o parte de ellas (Figura 1), de las preparaciones de las antenas y de la genitalia externa se realizó utilizando microscopio estereoscópico Nikon ZM10 y cámara fotográfica digital Nikon Coolpix p5100 con adaptador Nikon EH-62a. Los dibujos de las antenas y genitalia externa (Figuras 2 y 4) fueron realizados por los autores a mano, a partir de las fotos ampliadas de dichas estructuras, y posteriormente fueron editados y digitalizados. La agrupación de los juveniles en distintos estadios se realizó considerando el desarrollo de los rudimentos alares (Figura 1) y de la genitalia externa (Figura 2) de acuerdo a Uvarov (1966), dado que en general estas son características constantes en el desarrollo de las ninfas de los acridios (Turk y Barrera, 1979).
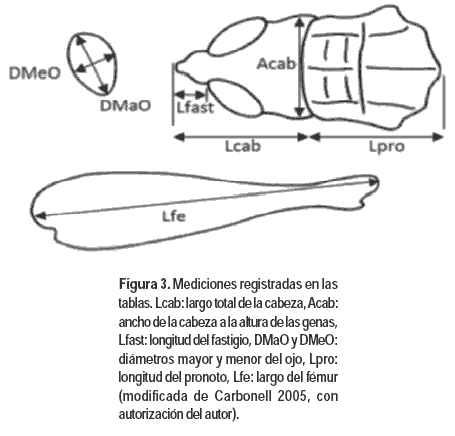
Con el objetivo de complementar la caracterización de los estadios ninfales con caracteres morfométricos y establecer si existen diferencias significativas a este nivel, se evaluaron diferentes medidas corporales: ancho de la cabeza a la altura de las genas, diámetro mayor y menor del ojo, y largo de la cabeza, del fastigio, del pronoto y del fémur posterior derecho (Figura 3). Para ello se tomaron fotos digitales de cada estructura a los efectos de realizar las mediciones correspondientes con el software Image Pro-Express 5. En total se midieron 20 ejemplares (10 de cada sexo) de las todos los estadios ninfales.
También se determinó el número de segmentos antenales de cada estadio ninfal. Para ello se contó el número de antenitos en seis ejemplares (tres de cada sexo) de cada estadio ninfal (Figura 4). Las antenas fueron aclaradas con KOH al 10 % y posteriormente lavadas con agua destilada. Para su montaje, se utilizó alcohol polivinílico.
Con toda la información de las variables morfológicas y morfométricas se realizó la descripción de los estadios.
Las variables morfométricas, fueron analizadas con modelos lineales generales (PROC GLM, SAS Institute, version 9.2), dado que la varianza fue homogénea y no se encontró asociación entre la media y la varianza. Las medias fueron comparadas utilizando la prueba de Tukey-Kramer (HSD) (P ≤ 0,05). El número de segmentos antenales, por ser una variable discreta con varianza heterogénea y con importante asociación entre la media y varianza, fue analizado utilizando modelos lineares generalizados con distribución Poisson (PROC GENMOD, SAS Institute, version 9.2). Los resultados son presentados como la máxima verosimilitud de la distribución Chi cuadrado. Para todas las variables el modelo consideró el estadio y el sexo como efectos fijos.
Resultados
Fueron colectados un total de 1221 individuos pertenecientes a B. bruneri. De acuerdo al análisis de los rudimentos alares, el desarrollo de la genitalia externa y de las antenas, el material colectado fue clasificado en cinco estadios ninfales.
Descripción general de las ninfas, caracteres diagnósticos
La coloración general del cuerpo varía del tostado o marrón amarillento al crema, a veces algo verdoso. Las manchas oscuras del cuerpo varían desde el marrón oscuro al ámbar oscuro. Desde el fastigio hasta el extremo abdominal, a los lados de la línea media del cuerpo se extienden dorsalmente dos bandas oscuras separadas por una banda de coloración clara. La extensión e intensidad de la coloración de estas bandas es variable según los estadios y los individuos. Es característico el patrón de coloración en X de las bandas oscuras en el pronoto y de las carenas laterales de coloración clara que lo acompañan, típico de los adultos, que se comienza a visualizar a partir del estadio II. Este patrón se torna cada vez más conspicuo a medida que avanza el desarrollo ninfal: la coloración de las carenas laterales vira desde el ámbar claro a blanco. En las caras laterales del pronto, en su porción anterior se observa una mancha oscura de forma casi circular, que comienza a ser visible a partir del estadio II.
Cabeza de perfil con el vértice marcadamente convexo, fastigio levemente convexo y frente casi recta por debajo de las antenas. Inclinación de la frente con respecto a la cabeza moderada (ángulo de aproximadamente 60 °), de forma similar al adulto. Las antenas varían de una forma achatada con su extremo apical ensanchado, en los primeros estadios, a filiformes en los siguientes estadios.
Las carenas laterales del pronoto, están interrumpidas entre el primer y segundo surco transversal, divergentes hacia la cabeza las anteriores y hacia el extremo caudal las posteriores. Las carenas laterales apenas prominentes en el estadio II se hacen cada vez más notorias, siendo las posteriores más anchas.
Fémures de las patas I y II ensanchados como en el adulto. Coloración del fémur posterior: en general uniforme, tibias: claras.
Descripción morfológica de los estadios
Estadio I:
Ninfas con banda dorsal media a lo largo del cuerpo de color claro, a los lados se extienden las bandas oscuras que se ensanchan levemente desde el fastigio hasta el extremo abdominal. Los rudimentos alares son imperceptibles (Figura 1 A). Las carenas laterales del pronoto ausentes. El borde posterior del pronoto es apenas escotado hacia la carena media. Las antenas están compuestas por 11 segmentos (Figura 4 A).
Genitalia externa:
♂: El noveno esternito abdominal (placa subgenital) posee una porción anterior de forma rectangular, más ancha que larga, y una porción posterior terminada en un margen libre, de bordes laterales subparalelos, con una escotadura profunda y semicurcular en el ápice y extremos aguzados (Figura 2 A). Cubre la porción basal de los paraproctos, sin llegar a alcanzar con los extremos la mitad de su longitud. Los paraproctos son visibles, alargados y de extremos redondeados. El epiprocto y los paraproctos son aproximadamente de igual longitud. Los cercos son triangulares con ápices acuminados, erectos.
♀: El octavo esternito abdominal es de forma rectangular con el ancho de aproximadamente doble longitud que el largo (Figura 2 F). El primer par de valvas (valvas ventrales) se visualiza como dos placas rudimentarias, en el borde distal de dicho esternito, unidas totalmente en la línea media y con los márgenes posteriores apenas redondeados. No llegan a sobrepasar el margen posterior del siguiente esternito. El tercer par de valvas (valvas dorsales) se origina en el borde posterior del noveno segmento abdominal y se observa como un lóbulo ancho, subdividido en dos lóbulos subtriangulares separados entre sí por una escotadura profunda en U invertida o en ángulo recto. No alcanzan a cubrir el tercio basal de los paraproctos. Los paraproctos son alargados y el epiprocto sobrepasa apenas en longitud a los paraproctos. Los cercos son similares a los de los machos.
Estadio II
Ninfas con bandas oscuras a los lados de la banda dorsal clara que extiende a lo largo del cuerpo. Comienza a hacerse aparente en el pronoto el diseño en X de las bandas oscuras. Se observan las carenas laterales del pronoto, de coloración clara y apenas prominentes. El borde posterior del pronoto tiene la misma forma que en el estadio anterior, con la escotadura en general más abierta. Las manchas oscuras en las caras laterales del pronto se hacen notorias en algunos ejemplares. Los rudimentos alares son visibles, apenas esbozados desde los extremos distales del meso y metanoto (Figura 1 B). Las antenas están compuestas de 11-12 segmentos en el macho y 11 en la hembra (Figura 4 B).
Genitalia externa:
♂: La porción anterior del noveno esternito y los cercos son similares a los del estadio anterior (Figura 2 B). La porción posterior se estrecha levemente hacia el ápice, la escotadura del extremo posterior es apenas cóncava. El extremo de la placa subgenital sobrepasa apenas la mitad de los paraproctos. Los epiproctos son apenas más pequeños que los paraproctos. Los cercos son de forma similar a la anterior categoría.
♀: La relación entre el ancho y el largo del octavo esternito es semejante a la del estadio anterior (Figura 2 G). El primer par de valvas está formado por dos lóbulos triangulares separados en su línea media, que cubren menos de la mitad basal del noveno esternito. El tercer par de valvas está formado por dos lóbulos triangulares, alargados, libres, unidos en la base por la línea media, apenas si llegan a cubrir la mitad de los paraproctos. El epiprocto es visible ventralmente y la relación de longitud con los paraproctos es similar a la del estadio I. Los cercos son similares al estadio anterior.
Estadio III
Ninfas con coloración similar al estadio anterior, excepto que hay una profundización en la pigmentación de las bandas oscuras en la zona del pronoto. El borde posterior del pronoto es casi recto. Las carenas están bien marcadas. Los rudimentos alares tienen el borde inferior agudo, y están orientados hacia atrás (Figura 1 C). Los machos y las hembras tienen 14-15, y 15 segmentos antenales respectivamente (Figura 4 C).
Genitalia externa:
♂: La placa subgenital es ancha y convexa, y se estrecha hacia el ápice, terminando en un extremo romo, sin escotadura (Figura 2 C) y sobrepasa la mitad de los paraproctos. La relación entre la longitud de los paraproctos y el epiprocto es similar a la del estadio II. Los cercos se visualizan sin mayores cambios de forma.
♀: El octavo esternito mantiene una forma similar a la de los estadios anteriores, sigue siendo más ancho que largo (Figura 2 H). El borde distal del mismo es recto o apenas cóncavo. El primer par de valvas tiene forma triangular, con ápices acuminados que superan el borde distal del noveno esternito abdominal. Estas cubren la base del tercer par de valvas, que son de forma triangular y alargada, y sobrepasan la mitad de los paraproctos. Comienzan a ser visibles las valvas medias o internas como dos pequeñas placas ovoideas alargadas, situadas entre las dorsales y las ventrales. El epiprocto y los paraproctos son de igual longitud. En los cercos no se observan mayores modificaciones.
Estadio IV
Coloración ninfal similar al estadio anterior. Carenas laterales del pronoto bien marcadas, las posteriores más anchas que las anteriores. Se visualizan claramente los surcos transversales del pronoto. El borde posterior del pronoto recto o levemente curvado hacia el extremo abdominal. Entre el estadio anterior y este, se produjo la torsión postero-dorsal, por lo que se observan los sacos alares invertidos, siendo los anteriores más largos que los posteriores (Figura 1 D). El número de segmentos antenales en los machos es de 18-19 y en las hembras de 19-20 (Figura 4 D).
Genitalia externa:
♂: La placa subgenital es alargada y se estrecha hacia el ápice. Tiene la superficie convexa, el extremo posterior incurvado dorsalmente y el borde distal convexo (Figura 2 D). El largo de la placa subgenital es apenas menor que el de los paraproctos, los cuales son apenas visibles a los lados. Los paraproctos y el epiprocto tienen la misma longitud. Los cercos se visualizan sin mayores cambos.
♀: El octavo esternito es de forma cuadrangular, con el ancho aproximadamente de igual longitud que el largo (Figura 2 I). El borde distal de este esternito es levemente sinuado con una proyección media aguzada, que insinúa el desarrollo de la futura guía de huevos entre el primer par de valvas. Estas son de forma triangular, con el extremo distal aguzado, y cubren la porción basal del tercer par de valvas. Del noveno segmento solo se visualizan pequeñas porciones laterales. Las valvas del tercer par son algo cónicas, cubren los paraproctos casi completamente hasta su extremo distal. Los paraproctos sobrepasan el extremo distal del epiprocto.Los cercos son similares al estadio anterior.
Estadio V
Coloración ninfal sin cambios. Carenas del pronoto bien marcadas y de color blanco. En el pronoto se observa un cambio en la forma del borde posterior, apenas se proyecta hacia atrás en ángulo obtuso romo. Los sacos alares están más desarrollados, ambos pares se disponen en tejado sobre el abdomen y cubren el tímpano ubicado en el primer segmento abdominal (Figura 1 E). Los machos tienen 20 segmentos antenales, mientras que las hembras tienen entre 20 y 21 (Figura 4 E).
Genitalia externa:
♂: La placa subgenital es de forma similar al estadio anterior, convexa y con el extremo incurvado dorsalmente. Se estrecha hacia el ápice, con el borde distal convexo y sobrepasa en longitud a los paraproctos (Figura 2 E). El epiprocto es más corto que los paraproctos. Los cercos permanecen sin mayores cambios de forma
♀: El octavo esternito, que formará la placa infragenital de la hembra adulta, es más largo que ancho. Presenta el margen posterior sinuado con dos concavidades próximas a la línea media y un proceso medio de forma triangular (guía de huevos) (Figura 2 J). El primer par de valvas es apenas menor en tamaño que el tercero. Las valvas del tercer par alcanzan en longitud el extremo abdominal, cubren totalmente los paraproctos y el epiprocto. El extremo de las valvas es redondeado y está apenas esclerificado. El epiprocto es un poco más corto que los paraproctos.Los cercos continúan sin mayores modificaciones.
Estudio morfométrico
Los resultados de los análisis de las variables consideradas indican que hubo diferencias altamente significativas entre los estadios (antenitos X2 = 28,55; gl = 4; P< 0,0001; largo de la cabeza F 4,90 = 356,49, P< 0,0001; ancho de la cabeza F 4,90 = 435,38, P< 0,0001; largo del fastigio F 4,90 = 128,69, P< 0,0001; diámetro mayor del ojo F 4,90 = 398,47, P< 0,0001; diámetro menor del ojo F 4,90 = 440,80, P< 0,0001; largo del pronoto F 4,90 = 798,80, P< 0,0001; largo del fémur F 4,90 = 845,06, P< 0,0001) y del sexo, con excepción del número de segmentos antenales que no presentó diferencias significativas (X2 = 0,03; gl = 4; P=0,8602), (largo de la cabeza F 4,90 = 89,74, P< 0,0001; ancho de la cabeza F 4,90 = 95,46, P< 0,0001; largo del fastigio F 4,90 = 25,74, P< 0,0001; diámetro mayor del ojo F 4,90 = 89,06, P< 0,0001; diámetro menor del ojo F 4,90 = 47,54, P< 0,0001; largo del pronoto F 4,90 = 798,80, P< 0,0001; largo del fémur F 4,90 = 82,83, P< 0,0001). Con excepción del número de segmentos antenales y de la longitud del fastigio, las restantes variables discriminaron a los cinco estadios dentro de cada sexo, dado que los valores medios registrados en cada una de ellas fueron significativamente diferentes entre sí (Cuadro 1). Al igual que en otras especies de acridios, las hembras fueron más grandes que los machos. Los análisis indican que en todos los estadios ambos sexos fueron diferentes en el largo de la cabeza, el diámetro mayor del ojo y el largo del fémur. Por otro lado las diferencias entre sexos en el ancho de la cabeza y el largo del pronoto se registraron a partir del estadio III (Cuadro 1). Respecto al diámetro menor del ojo, ambos sexos presentaron diferencias significativas en todos los estadios ninfales con excepción del estadio II.
Discusión
Borellia bruneri fue sin duda la especie dominante durante las explosiones poblacionales registradas en Uruguay en la década de los 50 en el siglo pasado, con densidades que superaron en más del doble a la segunda especie en importancia (Liebermann y Piran, 1941; Liebermann y Ruffinelli, 1946; Ruffinelli y Carbonell, 1953; Carbonell, 1957; Silveira Guido et al., 1958). Posteriormente su densidad poblacional disminuyó en forma tal que resultaba difícil de ser observada en el campo (Martinez Crosa, 2004). Esta característica fue un impedimento para avanzar en el conocimiento de su ciclo y de su fenología.
Este es el primer trabajo en el que se realiza una descripción de los cambios que ocurren durante el estado de ninfa de esta especie. Los resultados obtenidos permitieron caracterizar cinco estadios ninfales de B. bruneri. Los tres primeros correspondieron a individuos con los rudimentos alares del meso y metanoto ubicados lateralmente, mientras que en los dos últimos se observaron las tecas alares ya invertidas. Los cambios en la estructura externa de la genitalia de las hembras fueron más evidentes que los ocurridos en la genitalia externa de los machos. El estudio de la morfometría de los estados juveniles de los acridios es una herramienta frecuentemente utilizada para separar estadios ninfales (Turk y Barrera, 1976). Las variables morfométricas, largo de la cabeza, diámetro mayor del ojo y largo del fémur fueron diferentes entre los estadios y entre sexos. Estas medidas serían las más adecuadas para utilizar en estudios de estructuración de la población. Algunos autores plantean que en los diferentes estadios ninfales de varias especies de acridios ocurren diversos cambios en la estructura de la antena, en sus componentes, particularmente el número y tipo de segmentos del flagelo, por lo que la evaluación de los mismos es un método útil para separar estadios ninfales (Zolessi, 1956, 1958; Turk y Barrera, 1976; Santoro y Carames, 1973; Santoro, 1976; Santoro et al., 1978). Sin embargo, en este trabajo el número de segmentos antenales no diferenció a los distintos estadios. De todas las variables que discriminaron, la más adecuada para diferenciar en el campo los distintos estadios y los sexos, sin el auxilio de microscopio estereoscópico, fue el largo del fémur, por ser la estructura de mayor tamaño.
De acuerdo a la clasificación morfológica y el estudio morfométrico, esta especie tiene cinco estadios ninfales, tal como lo determinó Mariottini (2009) en Argentina. Silveira Guido et al. en 1958, cuando realizaron la cría de esta especie en el laboratorio sin condiciones controladas, determinaron que esta especie tenía seis estadios ninfales. Santoro (comunicación personal) mediante el análisis de las antenas de ejemplares adultos y de las antenas del primer estadio calculó la cantidad de estadios ninfales de esta especie en cinco.
En el manejo de estos insectos, ante una explosión poblacional es decisivo el momento de la prevención. Una de las herramientas fundamentales para la prevención, que colabora con la toma de decisiones correctas, es el seguimiento periódico y sistemático de las poblaciones, lo que permite determinar aproximadamente la estructura de la población en relación a los estadios ninfales (Berry et al., 1996). Esto es necesario para evitar pérdidas, dado que los tratamientos de control deben ser realizados cuando la mayor proporción de la población se encuentra en el tercer estadio de ninfa y no antes ni después de ese momento (Hewitt y Osanger, 1983). Así se asegura que la mayor cantidad de huevos hayan eclosionado y se evita la aplicación de medidas de control cuando aún se puede esperar eventos importantes de mortalidad por causas naturales en la población. Los tratamientos de control tampoco se deben retrasar porque a partir del cuarto estadio las ninfas son menos susceptibles a los insecticidas, el consumo de alimento aumenta significativamente y pueden producir daños. El control en estado adulto es injustificado porque para entonces ya se produjo el mayor daño La proporción de adultos no debería superar el 20 % (Hewitt, 1980; Hewitt y Osanger, 1983; Curvetto, 2009; Lorier et al., 2010).
Conclusiones
Fueron determinados cinco estadios ninfales distinguibles en base al desarrollo de los rudimentos alares y la genitalia externa. Las medidas del largo de la cabeza, diámetro mayor del ojo y largo del fémur posterior fueron los caracteres más útiles para separar los individuos de los distintos estadios que pertenecen al mismo sexo y entre sexos.
Agradecimientos
A Mabel Pessio y Eduardo García por su colaboración durante los muestreos.
Bibliografía
Bentancourt CM, Scatoni, IB, Morelli, E. 2009. Insectos del Uruguay. Montevideo: Facultad de Agronomía ; Facultad de Ciencias. 658p.
Berry JS, Onsager JA, Kemp WP, Mcnary T, Larsen J, Legg D, Lockwood JA, Foster RN. 1996. Assessing rangeland grasshopper populations. En: Cunningham GL, Sampson MW [Eds.]. Grasshopper integrated pest management user handbook. Washimgton : USDA. (Technical Bulletin ; 1809). pp. 1-12.
Carbonell CS. 1995. Revision of the tribe Scyllinini, Nov. (Acrididae: Gomphocerinae), with descriptions of new genera and species. Transactions of the American Entomology Society, 121: 87-152.
Carbonell CS. 1957. Vuelos en masa de acridoideos (Orthoptera) en el Uruguay. Revista de la Sociedad Uruguaya de Entomología, 2: 73 - 77.
Carbonell CS, Cigliano MM, Lange CE. 2006. Especies de Acridomorfos (Orthoptera) de Argentina y Uruguay [CD ROM]. La Plata : The Orthopterist´s Society at the Museo de La Plata. (Publications on Orthopteran Diversity).
Cigliano MM, Kemp WP, Kalaris T. 1995. Spatiotemporal characteristics of rangeland grasshopper (Orthoptera: Acrididae) regional outbreaks in Montana. Journal of Orthoptera Research, 4: 11 – 126.
COPR. 1982. The locust and grasshopper agricultural manual. London: Centre for Overseas Pest Research. 660p.
Curvetto R. 2009. La Tucura: una amenaza a la producción regional [En línea]. Consultado 28 setiembre 2014. Disponible en: http://inta.gob.ar/documentos/la-tucura-una-amenaza-a-la-produccion-regional/at_multi_download/file/1.%20tucura.pdf.
de Wysiecki MA, Arturi M, Torrusio S, Cigliano MM. 2011. Influence of weather variables and plant communities on grasshopper density in the Southern Pampas, Argentina. Journal of Insect Science, 11(109): 1 – 14.
de Wysiecki ML, Torrusio S, Cigliano MM. 2004. Caracterización de las comunidades de acridios (Orthoptera: Acridoidea) del partido de Benito Juárez, sudeste de la provincia de Buenos Aires, Argentina. Revista de la Sociedad Entomológica Argentina, 63(3-4): 87-96.
de Wysiecki ML, Sánchez N, Ricci S. 2000. Grassland and grasshopper community composition in northern La Pampa province, Argentina. Journal of Orthoptera Research, 9: 211-221.
Hewitt GB.1980. Plant phenology as a guide in timing grasshopper control efforts on Montana Rangelands. Journal of Range Management, 33 (4): 297 – 299.
Hewitt GB, Onsager JA. 1983. Control of grasshoppers on rangeland in the United States – a perspective. Journal of Range Management, 36: 202 – 207.
Joern A. 2000. What are the consecuences of non-linear ecological interactions for grasshopper control strategies? En: Lockwood JA, Latchininsky AV, Sergeev MG [Eds.]. Grasshoppers and Grassland Health : Managing Grasshopper Outbreaks without Risking Environmental Disaster. Boston: Kluwer Academic. pp. 131-143.
Liebermann J, Ruffinelli A. 1946. Catálogo de acridoideos uruguayos. Revista de la Asociación de Ingenieros Agrónomos, 74: 9 -21.
Liebermann J, Piran AA. 1941. Primera lista de acridios uruguayos. Montevideo: Dirección General y de Contralor de la Lucha Contra la Langosta. 12 p.
Lorier E, Zerbino S. 2009. Tucura: radiografía de una plaga. El País Agropecuario, 175: 36-38.
Lorier E, Miguel L, Zerbino MS. 2010. Manejo de tucuras. En: Altier N, Rebuffo M, Cabrera K. [Eds.] Enfermedades y plagas en pasturas. Montevideo: INIA. (Serie técnica; 183). pp. 51-71.
Mariottini Y. 2009. Biología y ecología de acridios (Orthoptera: Acridoidea) del Sur de la región Pampeana [Tesis de doctorado]. La Plata: Facultad de Ciencias Naturales y Museo. Universidad Nacional de La Plata. 203p.
Mariottini Y, de Wysiecki ML, Lange CE. 2013. Diversidad y distribución de acridios (Orthoptera: Acridoidea) en pastizales del Sur de la región Pampeana, Argentina. Revista de Biología Tropical, 61: 111-124.
Mariottini Y, de Wysiecki ML, Lange CE. 2012. Variación temporal de la riqueza, composición y densidad de acridios (Orthoptera: Acridoidea) en diferentes comunidades vegetales del Sur de la provincia de Buenos Aires. Revista de la Sociedad Entomológica Argentina, 71: 275-288.
Mariottini Y, de Wysiecki ML, Lange CE. 2011. Seasonal occurrence of life stages of grasshoppers (Orthoptera: Acridoidea) in the Southern Pampas, Argentina. Zoological Studies, 50: 737-744.
Martinez Crosa G. 2004. Parámetros comunitarios y hábitos alimentarios de un ensamble de acridomorfos (Orthoptera; Acridoidea) en una pradera natural [Tesis de maestría]. Montevideo: Facultad de Ciencias. Universidad de la República. 73 p.
Pfadt RE. 2002. Field guide to common western grasshoppers. 3a ed. Wyoming: Wyoming Agricultural Experiment Station. 46p. (Bulletin; 912).
Resh VH, Cardé RT. 2003. Orthoptera. En: Encyclopedia of Insects. San Diego: Academic Press. pp. 827-839.
Ruffinelli A, Carbonell CS. 1953. Segunda lista de insectos y otros artrópodos de importancia económica en el Uruguay. Revista de la Asociación de Ingenieros Agrónomos, 94: 33-82.
Sanchez N, de Wysiecki ML.1993. Abundancia y diversidad de Acridios (Orthoptera:Acrididae) en pasturas de la provincia de La Pampa, Argentina. Revista de Investigaciones Agropecuarias, 24: 29-39.
Santoro FH. 1976. Cálculo de la cantidad de estadios ninfales o de la edad por el análisis de la antena del adulto o de los dos estadios inmaturos previos, en diversas especies de acridios (Orthoptera – Acrididae). Revista de Investigaciones Agropecuarias, 12(1): 1-6.
Santoro FH, Carames A. 1973. Antenitos que determinan las variaciones cuantitativas postembrionales en la antena de doce especies de acridios de la Argentina (Insecta, Orthoptera). Revista de Investigaciones Agropecuarias, 10(4): 197-209.
Santoro FH, Uvagli A, Sisler GM de, Salto C, Carames A. 1978. Identificación de ninfas de siete especies del género Dichroplus (Orthoptera: Acrididae). Revista de Investigaciones Agropecuarias, 7(2): 89-112.
Silveira Guido A, Carbonell JF, Núñez O, Valdés E. 1958. Investigaciones sobre acridoideos en el Uruguay. Montevideo : Facultad de Agronomía. Universidad de la República. 485p.
Torrusio S, Cigliano MM, de Wysiecki ML. 2002. Grasshopper (Orthoptera: Acridoidea) and plant community relationships in the Argentine Pampas. Journal of Biogeography, 29: 221-229.
Turk S, Barrera M. 1979. Acridios del NOA III: Estudio bio-ecológico sobre siete especies del género Dichroplus. Stal (Orthoptera, Acrididae). Acta Zoológica Lilloana, 35(6): 785-805.
Turk S, Barrera M. 1976. Acridios del NOA I: Estudios biológicos, morfométricos y aspectos ecológicos de Chromacris speciosa (Thunberg) (Acrididae, Romaleinae). Acta Zoológica Lilloana, 32(6): 121-146.
Uvarov BP. 1966. I. Grassopers and locusts. London: Cambridge University Press. 481p.
Zolessi LC de. 1958. Bioecología y ontogenesis de Scotussa cliens (Stål) Lieb. (Acridoidea, Catantopidae) en el Uruguay. Montevideo: Facultad de Humanidades y Ciencias. Universidad de la República. 19p.
Zolessi LC de. 1956. Observaciones sobre Cornops aquaticum Br. (Acridoidea, Cyrtacanthacr.) en el Uruguay. Revista de la Sociedad Uruguaya de Entomología, 1: 3 - 28.

