










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Links relacionados
Compartilhar

 Permalink
PermalinkAgrociencia (Uruguay)
versão impressa ISSN 1510-0839versão On-line ISSN 2301-1548
Agrociencia Uruguay vol.17 no.2 Montevideo dez. 2013
Variables morfogenéticas y estructurales de cinco cultivares de Lolium sp.
Saldanha Sylvia 1, Cechini Anabel 2, Bentancur Oscar3
1Departamento de Producción Animal y Pasturas, Facultad de Agronomía, Universidad de la República. Estación Experimental Salto, Ruta 31, km 21,5, 50000 Salto, Uruguay. Correo electrónico: sylsal71@adinet.com.uy
2Departamento de Producción Animal y Pasturas, Facultad de Agronomía, Universidad de la República.
3Departamento de Biometría y Estadística, Facultad de Agronomía, Universidad de la República. Estación Experimental Mario A. Cassinoni, Ruta 3, km 363, 60000 Paysandú, Uruguay.
Recibido: 23/11//12 Aceptado: 24/10/13
Resumen
Se cuantificaron variables morfogenéticas: tasas foliares de elongación (TEF), senescencia (TSF), aparición foliar (TAF) y vida media foliar (VMF), y variables estructurales (número y largo de hojas por macollo y densidad de macollos) en cultivares de Lolium sp.: INIA Titán (Titán), INIA Cetus (Cetus), Conker (Conker), Maverick Gold (Maverick) y Dominó (Dominó). El objetivo fue evaluar si los cultivares diferían en estas variables, su respuesta a la temperatura y la utilidad de caracterizarlos para definir manejos apropiados para cada pastura. Se ajustaron modelos de regresión lineales de los flujos foliares con las temperaturas promedio diarias. Se evaluaron modelos cuadráticos cuando los modelos lineales no eran significativos. Se realizaron análisis de varianza de las variables para todo el período experimental (julio a setiembre) y las medias fueron separadas por el test de Tuckey (p≤0,05). En función del filocrón Conker y Maverick fueron materiales de alto recambio foliar, con altas TEF que permitirían alcanzar rápidamente el índice de área foliar (IAF) óptimo, con VMF menor y TSF mayores por lo que estos cultivares estarían más adaptados a defoliaciones frecuentes. Esto se asoció a una mayor densidad de macollos (no significativo estadísticamente). Los cultivares tetraploides Dominó y Titán se ubicaron en el otro extremo, con menor densidad de macollos, TAF, TEF y TSF y mayor VMF, por lo que se adaptarían a pastoreos menos frecuentes. Cetus presentó un comportamiento intermedio en las variables morfogenéticas.
Palabras clave: TASA DE APARICIÓN DE HOJAS, VIDA MEDIA FOLIAR, DENSIDAD DE MACOLLOS
Summary
Morphogenetic and Structural Variables in Five Cultivars of Lolium sp.
Morphogenetic variables: leaf elongation rate (TEF), leaf senescence rate (TSF), leaf appearance rate (TAF) and leaf lifespan (VMF), and structural variables (number and leaves length per tiller and tiller density) were estimated on cultivars of Lolium sp.: INIA Titán (Titán), INIA Cetus (Cetus), Conker (Conker), Maverick Gold (Maverick) and Dominó (Dominó). The objective was to evaluate if the cultivars were different in these variables, response to temperature, and the usefulness of characterizing each cultivar, in order to define the appropriate management for each sward. Lineal regression models were adjusted between leaf tissues fluxes and daily mean temperature. Quadratics´ models were evaluated, if showed no significant lineal regression. Analysis of variance was made for the different variables for the experimental period (July to September) and means were compared to Tuckey´s test (p≤0.05). According the phyllochron, Conker and Maverick were cultivars with rapid leaf turnover, with high TEF, that would reach quickly the optimal leaf area index (LAI), with a lower VMF and a higher TSF. This was associated with a higher tiller density (not statistically significant) and therefore these cultivars would be more adapted to frequent defoliation. Tetraploid cultivars Dominó and Titán stood at the other end, with a lower density of tillers, TAF, TEF y TSF, and a higher VMF, so they would be adapted to less frequent grazing. Cetus presented an intermediate behaviour for morphogenetic variables.
Keywords: LEAF APPEARANCE RATE, LEAF LIFESPAN, TILLER DENSITY
Introducción
El crecimiento aéreo de las plantas resulta de un programa de morfogénesis definido por las demandas de recursos provistos por la intercepción lumínica y su suministro a través de la fotosíntesis (Lemaire y Agnusdei, 2000; Gastal et al., 2004; Lemaire y Chapman, 1996). Este crecimiento aéreo puede describirse y analizarse a partir de los mecanismos morfogenéticos de aparición y expansión de nuevos órganos en la planta y su desaparición por senescencia (Lemaire y Agnusdei, 2000). En pasturas de tipo templado y en crecimiento vegetativo, la morfogénesis es función de tres características principales: tasa de elongación foliar (TEF), tasa de aparición de hojas (TAF) y la vida media foliar (VMF) (duración de vida de la hoja). Estas características morfogenéticas determinan las condiciones de estructura de la pastura, como ser número de hojas por macollo, ancho y largo de láminas (tamaño foliar) y número de macollos (Chapman y Lemaire, 1993; Lemaire y Chapman, 1996).
Una característica importante de las gramíneas es el rápido recambio de los tejidos, a través de la producción continua de nuevas hojas y la muerte de las hojas más viejas. El tejido foliar removido por el pastoreo animal es principalmente determinado por la tasa de producción de nuevas hojas y la tasa de senescencia foliar (TSF), las cuales son determinadas por la actividad morfogenética de las plantas (Chapman y Lemaire, 1993).
Conocer las variables morfogenéticas y cómo las mismas son afectadas por las condiciones del ambiente permite deducir los manejos más adecuados para cada genotipo como: el período entre pastoreos (en función de la VMF y el número máximo de hojas / planta), la densidad de siembra (en función de la tasa de macollaje) o el método de pastoreo (rotativo o continuo), entre otros.
El género Lolium sp. (raigras) posee por su aptitud forrajera varias especies cultivadas y naturalizadas en regiones templadas de todo el mundo (Casler y Kallenbach, 2007). En nuestro país se ha ampliado notablemente la oferta de materiales genéticos forrajeros (cultivares) de este género y seguramente continuará aumentando. La evaluación nacional de los cultivares se realiza con cortes simultáneos a todos los materiales para poder comparar la producción de forraje, es decir se aplica un manejo de defoliación común, lo que limita la manifestación de las diferencias productivas entre los cultivares.
Es importante conocer si existen diferencias morfo-fisiológicas entre cultivares que justifiquen distintas condiciones ambientales de evaluación para cada material (nivel de N, frecuencia de cortes, altura de corte, localización geográfica).
En la Estación Experimental de la Facultad de Agronomía en Salto (EEFAS) de la Universidad de la República, se sembraron cinco cultivares de Lolium sp.: INIA Titán (Titán), INIA Cetus (Cetus), Conker (Conker), Maverick Gold (Maverick) y Dominó (Dominó), con el objetivo de: 1. cuantificar variables morfogenéticas (TEF, TSF, TAF y VMF) y estructurales (número de hojas por macollo, largo y ancho de láminas) en los cultivares, 2. conocer la relación de los flujos de tejidos foliares con la temperatura, 3. evaluar la utilidad de definir características estructurales (morfo-métricas) como complemento en la caracterización de un cultivar forrajero, insumo necesario para realizar recomendaciones de manejo adecuadas para cada cultivar.
Materiales y métodos
Condiciones experimentales, manejo del ensayo, tratamientos y diseño experimental
Se instaló el experimento en la EEFAS (31º25‘S; 57º55‘W) en un Brunosol éutrico profundo de la Unidad Itapebí-Tres Árboles, correspondiente a la formación Arapey. Se sembró en líneas a 15 cm, 400 semillas viables/m2 de cada material el 10 de marzo, en tierra laboreada y fertilizada con 63 unidades de P2O5 y 60 unidades de N. Se refertilizó el 4 de julio con 30 kg de N/ha. Los tratamientos fueron los cinco cultivares de raigrás: Maverick, Conker, Dominó, Cetus y Titán.
El diseño fue en bloques completos al azar con dos repeticiones, las parcelas eran de 5 m x 3 m.
Estimación de las variables
Las determinaciones morfométricas se realizaron con una frecuencia semanal durante tres meses (desde el 5 de julio al 25 de setiembre) en 16 macollos de cada material, identificándolos con un anillo de alambre fino de diferentes colores. Se midió la longitud de la lámina verde de cada hoja (las que se enumeraban por edad) con una regla milimétrica, diferenciando el tipo de hoja: en elongación (distancia desde el ápice emergente hasta la lígula de la hoja anterior), o expandida (distancia desde la lígula hasta el extremo distal de la lámina o hasta el borde del área senescente), indicándose si estaba cortada.
Flujos de tejidos
Crecimiento: la TEF en mm/macollo/día se calculó como la sumatoria de las variaciones positivas en el largo foliar de cada lámina en crecimiento entre dos mediciones sucesivas. Senescencia: la TSF en mm/macollo/día se calculó como la sumatoria por macollo de las variaciones negativas en largo foliar de las láminas entre dos medidas sucesivas. Crecimiento neto del forraje: la determinación de TNF se realizó en base a la diferencia entre la TEF y la TSF diaria. Tasa de aparición foliar: la TAF se estimó como las hojas aparecidas por macollo por día, promedio de los macollos de cada parcela, entre cada medición sucesiva. Con la temperatura promedio acumulada entre dos mediciones sucesivas (temperatura base de 0 ºC) se determinaron las TEF, TSF, TNF y TAF en mm/macollo/ºC.
Intervalo de aparición foliar: el intervalo de aparición foliar (promedio de días entre la aparición de dos hojas sucesivas) se calculó como el inverso de la TAF. Con este dato y la temperatura promedio acumulada durante esos días se obtuvo el filocrón (intervalo de aparición foliar en grados día de crecimiento). Vida media foliar: la VMF se estimó a partir del producto del intervalo de aparición de hojas y el número de hojas vivas por macollo (Chapman y Lemaire, 1993).
Características estructurales estimadas semanalmente
El largo total de láminas verdes por macollo se determinó a través de la suma de los largos de todas las láminas. Es una medida indirecta del área fotosintética por macollo. También se evaluó el largo promedio de lámina expandida sin senescencia ni defoliación, medida indirecta del tamaño foliar, el número total de hojas por macollo, número de hojas en elongación y el número de hojas ya expandidas.
Características estructurales estimadas con muestreos destructivos
El 21 de junio y el 3 de agosto se cortó a ras del suelo con tijera manual una muestra por parcela de 0,2 x 0,5 m. Se separaron visualmente los macollos por tamaño en tres categorías: grandes, medios y pequeños, contabilizándolos y pesándolos luego de su secado hasta peso constante, para estimar la densidad de macollos por metro cuadrado, la composición de estos según tamaño y el tamaño promedio de macollo para cada cultivar en biomasa seca.
Análisis estadístico
Se estudiaron las medias de cada variable por cultivar (análisis de varianza) para todo el período experimental. Las medias de los efectos significativos fueron separadas usando el test de Tuckey (p≤0,05). Si la probabilidad era mayor a 0,05 y menor a 0,10 se separaron las medias de los cultivares por la prueba de LSD Fisher al 10%. El modelo utilizado fue:
Yij = μ + ζi i + bj + eij
siendo Y = variable, μ = la media poblacional, ζi = el efecto del cultivar, b = el efecto bloque y e= el error experimental.
Se ajustaron modelos de regresión lineal entre las variables TEF, TSF, TNF, TAF y la temperatura diaria acumulada. Luego se compararon los límites superior e inferior de los parámetros de las ecuaciones lineales para determinar si las funciones diferían o no entre cultivares. En el caso de que no existiera un ajuste lineal significativo, se evaluaron modelos de regresión cuadráticos. El criterio de significación utilizado en los ajustes de modelos de regresión fue de 5%. Se realizó un análisis de componentes principales con todas las variables, con el objetivo de corroborar si las asociaciones entre los cultivares, y entre las variables y los cultivares, son las mismas que las obtenidas al analizar individualmente estas. Los análisis se realizaron con el programa estadístico INFOSTAT versión libre.
Resultados
TEF de los cultivares y su relación con la suma térmica
La TEF foliar media de los distintos cultivares para el período experimental por día o por grado centígrado día fue significativa (Cuadro 1), siendo mayor en Conker que en Titán.
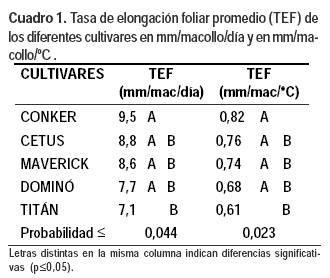
La suma térmica explicó significativamente (p<0,0001) el 95% de las variaciones observadas en las TEF por macollo de todos los cultivares, ajustando a un modelo lineal (Cuadro 2).
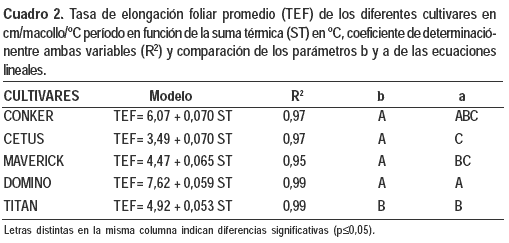
Comparando los parámetros de las ecuaciones lineales obtenidas se observó que el incremento en centímetros de hoja/macollo por cada grado centígrado de temperatura acumulada fue menor en Titán que en los otros cultivares.
TSF de los cultivares y su relación con la suma térmica
Las diferencias fueron significativas (p≤ 0,0002) entre cultivares para esta variable. La TSF por macollo y por día o por grado centígrado de Conker y Maverick fue en valor 84% mayor que la tasa de Dominó y Titán (Cuadro 3). La TSF de Cetus fue intermedia. Esta respuesta diferencial se reflejó en los patrones de acumulación neta de tejido foliar.
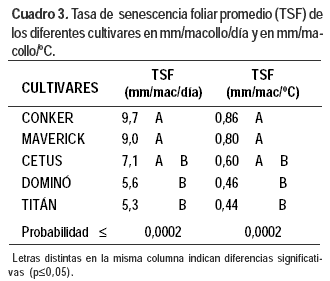
La suma térmica en el modelo lineal explicó el 92% de las variaciones observadas en las TSF de todos los cultivares, con un nivel significancia p≤ 0,0001 (Cuadro 4).
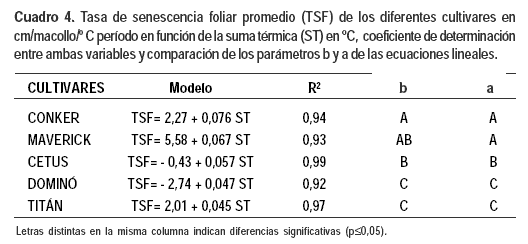
Comparando las relaciones entre la suma térmica con la TSF por macollo de todos los cultivares, el incremento de esta tasa con la temperatura fue similar en Titán y Dominó y menor al de los otros cultivares. Conker fue el cultivar que presentó mayor incremento de la TSF con la temperatura.
TNF de los cultivares y su relación con la suma térmica
Los cultivares no difirieron en la TNF por macollo por día y/o por grado centígrado. En Maverick y Conker fue negativa. Este último es el material de mayor TEF pero al ser la TSF superior, la TNF fue negativa. Seguramente en estos materiales debió realizarse un corte durante el período experimental (Cuadro 5).
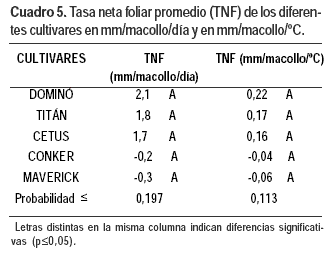
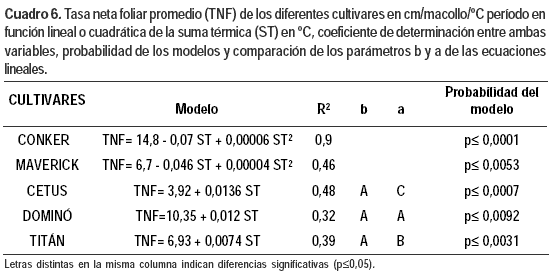
El ajuste lineal de la TNF con la temperatura fue significativo en: Titán, (p≤ 0,0031), Cetus (p≤0,0007) y Dominó (p≤ 0,0092), pero con coeficientes de determinación entre ambas variables medios a bajos. En Conker (p≤0,0001) y Maverick (p≤0,0016) el ajuste fue a un modelo cuadrático (Cuadro 6). Los modelos lineales no se diferenciaron en el coeficiente de regresión pero sí en la ordenada en el origen.
TAF de los cultivares y su relación con la suma térmica
Si bien no se detectaron diferencias significativas entre cultivares en la TAF por día, el filocrón varió entre los materiales evaluados (Cuadro 7). La suma térmica en el modelo lineal explicó significativamente (p≤0,0001) desde el 95% hasta el 99% de las variaciones observadas en la TAF (Cuadro 8).
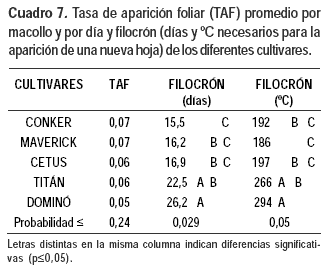
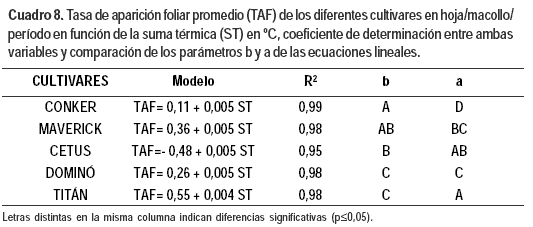
El incremento de la TAF con la temperatura fue diferente según el cultivar; fue mayor en Conker que en Cetus y en Dominó y Titán fue menor que en los otros cultivares.
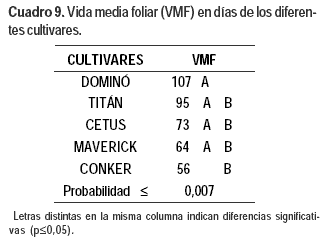
VMF de los cultivares
El cultivar Conker, tuvo menor VMF que Dominó (Cuadro 9).
Largo total de láminas verdes por macollo y largo promedio de lámina de una hoja completamente elongada, sin senescencia y sin defoliación, promedio para todo el período
Los cultivares se diferenciaron significativamente (p≤ 0,0001) en el largo total de hojas verdes (Cuadro 10). El largo total de hojas verdes por macollo en Conker y Cetus fue mayor que en Dominó y Titán, lo que era esperable ya que ambos presentaron mayores valores de TEF.

El largo promedio de la lámina de las hojas ya expandidas sin senescencia y sin defoliación mostró diferencias significativas (p≤ 0,067) para los diferentes cultivares, presentando Conker láminas más largas que Maverick, Dominó y Titán.
Número de hojas por macollo promedio para todo el período
La variable número de hojas totales fue significativa (p≤ 0,0001) para los cultivares en evaluación. Conker y luego Maverick fueron los cultivares con menor número de hojas totales. Las diferencias entre cultivares se deben principalmente al número de hojas ya expandidas. El número de hojas totales por macollo para cada cultivar no varió con las fechas a excepción de Conker (Cuadro 11).
Características estructurales medidas en dos momentos
Únicamente en la primera fecha y en dos variables se detectaron diferencias entre materiales (Cuadro 12).
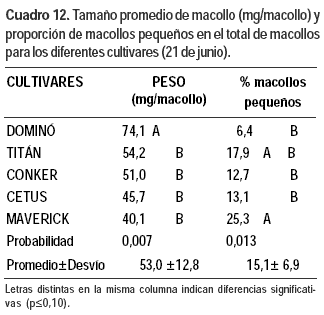
El peso seco de macollo (mg) promedio de los cultivares fue de 53,0 ± 12,8. El cultivar Dominó tuvo mayor (55%) peso promedio por macollo que los demás cultivares y baja proporción de macollos pequeños.
Las otras variables no presentaron diferencias significativas entre cultivares y no existió interacción fecha cultivar. La variación de los valores fue alta e importante biológicamente, lo que indicaría la necesidad de un mayor número de muestras para estimarlas. Por esto se presenta en el Cuadro 13 los valores para cada cultivar (promedio de ambas mediciones). La densidad de macollos por metro cuadrado promedio de todos los cultivares fue de 2127 ± 563, con una proporción de macollos grandes de 30 ± 16%, y de macollos chicos de 21,8 ± 10%.
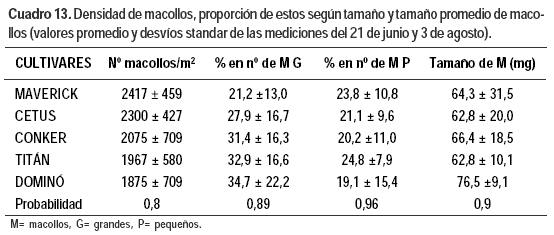
Los cultivares con mayor número de macollos presentaron mayor proporción de macollos pequeños y el peso promedio de los macollos fue menor. Una excepción a lo esperable fue el cultivar Titán en que a pesar de su baja densidad de macollos, estos fueron de bajo peso.
Análisis de componentes principales con características morfométricas y estructurales
El estudio multivariado nos permite el análisis en forma conjunta de las variables. Como se observa en la Figura 1, el 57% de la variabilidad total está explicada por el componente 1 que diferencia claramente los cultivares Dominó y Titán de los otros materiales, principalmente de Conker y Maverick. Las variables de mayor peso en este componente son TNF (0,37), proporción de macollos grandes (0,36), VMF (0,34), y tamaño de macollo (0,33). El componente 2 explica un 29% de la variación y distingue principalmente a Conker de Maverick, siendo importantes las variables largo total de láminas verdes por macollo (-0,45), número de hojas expandidas (0,45) y largo máximo de lámina expandida sin senescencia ni defoliación (0,40).
Las variables estudiadas distinguen a Dominó y Titán, cultivares tetraploides, de los otros materiales diploides. No se diferencian de acuerdo a si el cultivar es Lolium multiflorum tipo multiflorum (Dominó, Titán y Conker) o tipo westwerwoldicum (Cetus). Maverick es un cultivar de Lolium hybridum que surge del cruzamiento entre Lolium perenne y Lolium multiflorum.
Discusión
Flujos de tejidos foliares y sus relaciones con la suma térmica
El ajuste lineal de la TEF de los cultivares con la suma térmica coincide con resultados de otros autores cuando las temperaturas son mayores a 8 ºC (Gastal et al., 1992). En este trabajo, solo en un período se registró una temperatura promedio inferior (6,5 °C). Para la mayoría de las especies templadas la respuesta de la TEF a la temperatura promedio diaria es lineal entre 12 °C y 20-25 °C (Lemaire y Chapman, 1996). Las materiales de raigrás presentaron similar eficiencia de elongación (coeficiente b) por grado centígrado de temperatura diaria acumulada, a excepción de TITAN, y difirieron a nivel de ordenadas en el origen, interpretándose este parámetro como indicador de la capacidad para crecer a bajas temperaturas (Agnusdei et al., 1996).
La TEF promedio para el período estudiado fue mayor en Conker que en Titán. La mayor TEF de Conker determinó un mayor tamaño de hoja ya expandida y un mayor largo total de láminas verdes por macollo. Una rápida expansión de área foliar es un rasgo deseable principalmente en las primeras etapas de crecimiento de los cultivos, ya que lleva a un rápido cierre del canopeo reduciendo la evaporación de la superficie del suelo y aumentando el uso eficiente del agua (Richards et al. citados por Bultynck et al., 2004).También conlleva a que sea más competitivo con las malezas por la intercepción de luz (Lemaire, 2001). Altas TEF producen rápidamente una mayor superficie fotosintética y permiten promover una alta tasa de establecimiento y de rebrote después del pastoreo (Gao y Wilman, 1994). Si bien el cultivar Conker tuvo alta TEF, esto no se correlacionó negativamente con el número de macollos, lo cual era esperable para ciertos autores tal como Casal et al. (1985), Skinner y Nelson (1994) y Nelson (2000). La falta de correlación entre la TEF y la densidad de macollas del cultivar Conker se explica porque este cultivar no alargó el filocrón sino que lo acortó.
Tampoco tuvo el mayor rendimiento de forraje y rendimiento por macollo, que son rasgos esperables para un cultivar de alta TEF y de hojas largas. Fue el cultivar que, junto a Maverick, presentó mayor incremento de la TSF con la temperatura. La TSF de ambos fue un 84% mayor que la de Dominó y Titán. Cetus tuvo en valor intermedio.
Esta respuesta se reflejó en los patrones de acumulación neta de tejido foliar. Sin embargo esta variable no fue significativa entre cultivares. De todos modos la TNF por macollo por día y/o por grado centígrado de Maverick y Conker fue menor, e incluso negativa, lo que se relaciona con su alta TSF. La acumulación de material senescente estaría afectando la capacidad fotosintética y por ende la productividad. A los pocos días de iniciado el estudio estos cultivares posiblemente ya habían alcanzado su IAF óptimo. La tasa de crecimiento de la pastura aumenta hasta que el 95% de luz incidente es interceptada, siendo la acumulación neta de forraje máxima. Luego la TNF comienza a disminuir porque las hojas en las capas inferiores del canopeo están a la sombra, debajo de su punto de compensación de luz, lo que resulta en una pérdida de carbono neto a través de estas hojas por respiración (Chapman y Lemaire, 1993). A esto se podría sumar el hecho de que las hojas senescentes en invierno son las producidas en otoño, y por lo tanto de mayor tamaño foliar que las nuevas hojas, lo que determina menores tasas netas de acumulación de forraje (Carámbula, 2002a). Esto explica el ajuste cuadrático de la TNF en Conker y Maverick, que estaría indicando la necesidad de haber realizado un corte durante el período experimental al menos en estos materiales. Posteriormente al aumentar el IAF óptimo con la intensidad de luz (Chapman y Lemaire, 1993; Lemaire y Chapman, 1996) la TNF de estos cultivares retomó valores positivos a partir del 18 de agosto.
En función del filocrón podríamos concluir que Conker, Maverick y Cetus serían cultivares de alto recambio foliar, con alta densidad de macollos que garantizan un buen rebrote después de la defoliación. La alta TAF indica que deberían ser defoliados con mayor frecuencia (cada 56 a 73 días) para evitar que se produzcan pérdidas de material por senescencia, ya que ese rápido recambio implica una rápida restitución de área foliar y una tasa de macollaje potencial mayor, caracteres que confieren a las plantas aptitudes para adaptarse a una extracción más frecuente de tejido foliar. Ambas variables, la TSF y la TAF, están relacionadas (Colabelli et al., 1998).
Los otros cultivares alcanzarían su IAF optimo más tarde en el tiempo. Dominó y Titán presentaron los mayores filocrones (significativo al 10%) y la menor TEF, lo cual resulta en un crecimiento más lento, y al ser la TSF menor, determina un lento recambio foliar, características estas asociadas a los materiales tetraploides que poseen ciclos más largos (Carámbula, 2002b). La capacidad de rebrote después de la defoliación puede ser menor por una menor eficiencia fotosintética del área foliar remanente y por un menor número de meristemos, ya que la densidad de macollos es menor. Al tender a ser menor la TAF, la TSF sería menor, lo que concuerda con los datos obtenidos, donde la máxima acumulación neta de tejido foliar se dio más tardíamente en el tiempo. Estos genotipos serían aptos para sistemas de utilización que involucren defoliaciones menos frecuentes (cada 95 a 107 días) por su menor resistencia al pastoreo.
Al igual que otros autores (Chapman y Lemaire, 1993; Frank y Bauer, 1995; Kirby, 1995; Wilhem y McMaster, 1995; Colabelli et al., 1998) se obtuvieron altos coeficientes de determinación para las relaciones lineales entre el número de hojas producidas en un determinado intervalo de tiempo y la temperatura acumulada, confirmando que esta última es un factor que controla la TAF. En la mayoría de las especies C3 la respuesta de la TAF a la temperatura es lineal (Lemaire y Agnusdei, 2000). El incremento de TAF con la temperatura fue variable según el cultivar como se discutió anteriormente. Fue mayor en Conker que en Cetus y en este a su vez fue mayor que en Dominó y Titán.
VMF
La VMF puede ser utilizada para caracterizar la acumulación de los tejidos foliares verdes. Determina el lapso de tiempo para alcanzar el máximo de número de hojas vivas que un macollo individual puede soportar cuando su rendimiento techo es alcanzado (Lemaire y Agnusdei, 2000). Nos permite saber cuál es el tiempo de rebrote máximo que hay respetar antes de que se inicien pérdidas por senescencia.
Conker tuvo menor VMF que Dominó, lo que confirma su rápido recambio foliar, explicado por su mayor TSF, su menor filocrón en días y la menor cantidad de hojas vivas por macollo, comparado con el resto de los cultivares. En función de la VMF Dominó tendría el mayor rendimiento techo y Conker el menor. Esto explica también por qué durante el período experimental, al no realizarse cortes durante el mismo, Conker y Maverick presentaron TNF negativas. Cuando el intervalo de defoliación es menor que la VMF, solo la fracción de material de la hoja debajo de la altura de la defoliación tiene senescencia y se descompone, pero cuando el intervalo de defoliación es mayor que la VMF, una mayor proporción del material foliar se pierde por senescencia y la diferencia entre producción primaria y producción cosechable aumenta (Chapman y Lemaire, 1993). Según este trabajo, Conker permitiría un manejo de defoliaciones más frecuentes para evitar pérdidas por senescencia y por el contrario Dominó se adaptaría a un manejo de defoliaciones menos frecuentes.
Características estructurales
Conker y Cetus presentaron mayor largo total de hojas verdes por macollo, resultado esperable dadas las TEF mayores. Sin embargo ambos difieren en el número de hojas totales. Conker presenta menor número de hojas implicando láminas más largas. El número de hojas vivas totales por macollo está determinado por el producto entre la VMF y la TAF (Chapman y Lemaire, 1993).
La combinación de la TEF y la TAF permite determinar la longitud foliar de las hojas completamente expandidas (longitud foliar final) o sea el tamaño foliar (Lemaire y Chapman, 1996). Conker tuvo el menor intervalo de aparición de hojas expresado en días y fue el cultivar de mayor tamaño foliar, lo que no concuerda con Lemaire y Chapman (1996) para quienes el tamaño foliar es inversamente proporcional a la TAF.
La alta TEF por macollo de Cetus estaría determinada por un mayor número de hojas (número de meristemas intercalares) creciendo activamente. Los macollos con mayor proporción de hojas en crecimiento y con alta capacidad de crecimiento por hoja, podrían hacer un menor uso de los sitios de macollaje (Berone et al., 2007). En el caso de Cetus y Conker no sucedió de este modo porque la densidad de macollos fue alta. La falta de asociación negativa entre tamaño foliar y densidad de macollos concuerda con los resultados de Bahamani et al. (2000), en los que la selección por TEF altas y longitud de láminas largas se asocia a un sitio de llenado reducido, pero no necesariamente resulta en un menor número de macollos por planta en ciertas especies.
Los cultivares tetraploides evaluados, Titán y Dominó, presentaron menor cantidad de macollos por planta que los diploides, lo cual era previsible. En Dominó se dio una relación inversa entre número de macollos y tamaño de los mismos (Mathew et al., 1995) ya que el peso promedio por macollo el 21 de junio fue mayor (155%) al de los demás cultivares y fue menor la proporción de macollos chicos (37% del promedio de los demás cultivares). Titán, a pesar de su baja densidad de macollos, presentó macollos de bajo peso. Seguramente el nitrógeno presente en el suelo fue escaso y afectó la expresión del potencial de este cultivar, poco adaptado a bajos niveles de fertilidad (Carámbula, 2002b).
Maverick mostró alta densidad de macollos y alta proporción de macollos chicos, lo que se asocia con su tendencia a presentar una alta TAF, ya que esta determina directamente el número potencial de macollas (Chapman y Lemaire, 1996).
Las diferencias morfogenéticas y estructurales observadas sugieren contrastes inter-varietales marcados en los tiempos de máxima acumulación y recomposición del tejido foliar, en el potencial de macollaje, y por ende en la adaptación de las especies a diferentes regímenes de defoliación. Estas variables deberían considerarse en la elaboración de estrategias eficientes y sostenibles de utilización del forraje y al evaluar dichos materiales.
Bibliografía
Agnusdei MG, Colabelli M, Mazzanti A. 1996. Morfogénesis de especies nativas y naturalizadas de la pampa deprimida (Argentina). Revista Argentina de Producción Animal, 16 (1): 245-246.
Bahmani J, Hazard l, Varlet – Grancher C, Betin M, Lemaire G, Matthew C, Thom E. 2000. Differences in tillering of long and short leaved perennial ryegrass genotypes under full light and shade treatments. Crop Science, 40: 1095-1102.
Berone GD, Lattanzi FA, Colabelli MR, Agnusdei M. 2007. Análisis del crecimiento por macollo en dos cultivares de Bromus stamineus y dos de Lolium perenne. Revista Argentina de Producción Animal, 27(1): 203-204.
Bultynck L, Ter Streege MW, Schortemeyer M, Poot P, Lambers H. 2004 From individual leaf elongation to whole shoot leaf area expansion: a comparison of three aegilops and two Triticum species. Annals of Botany, 94: 99-108.
Carámbula M 2002a. Principios básicos de la producción de pasturas y forrajes. En: Pasturas y Forrajes Potenciales y alternativas para producir forraje. Tomo I. Montevideo: Editorial Hemisferio Sur. pp. 41-82.
Carámbula M 2002b. Las gramíneas. En: Pasturas y Forrajes Potenciales y alternativas para producir forraje. Tomo I. Montevideo: Editorial Hemisferio Sur. pp. 85-96.
Casal JJ, Deregibus VA, Sanchez RA. 1985. Variation in tiller dynamics and morphology in Lolium multiflorum Lam. vegetative and reproductive plant as affected by differences in red/far red irradiation. Annals of Botany, 56: 553-559.
Casler MD, Kallenbach RL. 2007. Cool season grasses for humid areas. En: Barnes R, Nelson J, Moore K, Collins M. [Eds.]. Forages The Science of grassland agriculture. Vol.II. Iowa: Blackwell Publishing. pp. 211-213.
Chapman DF, Lemaire G. 1993. Morphogenetic and structural determinants of plant regrowth after defoliation En: Baker MJ [Ed.]. Grassland of Our World. Wellington: SIR Publishing. pp. 95-104.
Colabelli M, Agnusdei M, Mazzanti A, Labreveux M. 1998. El proceso de crecimiento y desarrollo de gramíneas forrajeras como base para el manejo de la defoliación [En línea]. Boletín Técnico (INTA Balcarce), 148: 1 -14. Consultado 3 agosto 2011. Disponible en: http://www.vet.unicen.edu.ar/html/Areas/Introduccion%20a%20los%20Sistemas%20Prod/Documento/2009/LecRecom/crecimiento%20y%20desarrollo%20de%20GRAM%C3%8DNEAS.pdf.
Frank AB, Bauer A. 1995. Phyllochron Differences in Wheat, Barley, and Forage Grasses. Crop Science, 35: 19-23.
Gao Y, Wilman D. 1994. Leaf development in eight related grasses. Journal of Agricultural Science, 123: 41-46.
Gastal F, Lemaire G, Lestienne F. 2004. Defoliation, shoot plasticity, sward structure and herbage utilisation [Cd-Rom]. En: II Symposium in Grassland. Ecophysiology and Grazing Ecology; 11-14 octubre; 2004; Curitiba, Paraná, Brasil.
Gastal F, Belanger G, Lemaire G. 1992. A model of the leaf extension rate of tall fescue in response to nitrogen and temperature. Annals of Botany, 70: 437- 442.
Kirby EJM. 1995. Factors Affecting Rate of Leaf Emergence in Barley and Wheat. Crop Science, 35: 11-19.
Lemaire G. 2001. Ecophysiology of Grassland: Dynamic Aspects of Forage Plant Populations in Grazed Swards [Cd-Rom]. En: Proceedings XIX International Grassland Congress; 11- 21 febrery; 2001; Sao Paulo, Brasil. 25 p.
Lemaire G, Agnusdei M. 2000. Leaf tissue turnover and efficiency of herbage utilization. En: Lemaire G, Hodgson J, De Moraes A, Nabinger C, Carvalho PC de F. [Eds.]. Grassland Ecophysiology and Grazing Ecology. Oxford : CABI Publishing. pp. 265-287.
Lemaire G, Chapman D. 1996. Tissue flows in grazed plant communities. en: Hodgson J, Illius AW. [Eds.].The Ecology and Management of Grazing Systems. Wallingford: CAB International. pp. 3-35.
Matthew C, Lemaire G, Sackville Halmiton N R, Hernández - Garay A. 1995. A modified self-thinning equation to describe size/density relationships for defoliated swards. Annals of Botany, 76: 579-587.
Nelson CJ. 2000. Shoot morphological plasticity of grasses: leaf growth vs. tillering. En: Lemaire G, Hodgson J, De Moraes A, Nabinger C, Carvalho PC de F. [Eds.]. Grassland Ecophysiology and Grazing Ecology. Oxford: CABI Publishing,. pp. 101-126.
Skinner RH, Nelson CJ. 1994. Role of leaf appearance rate and the coleoptile tiller in regulating tiller production. Crop Science, 34: 71 – 75.
Wilhelm WW, McMaster G. 1995. Symposium on the phyllochron. Importance of the phyllochron in studying development and growth in grasses. Crop. Science, 35: 1-3.

