










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Links relacionados
Compartilhar

 Permalink
PermalinkAgrociencia (Uruguay)
versão impressa ISSN 1510-0839versão On-line ISSN 2301-1548
Agrociencia Uruguay vol.16 no.2 Montevideo dez. 2012
Respuesta poblacional invernal de dos gramíneas nativas a la fertilización nitrogenada y oferta de forraje
Zanoniani Ramiro1, Boggiano Pablo1, Cadenazzi Mónica1
1Facultad de Agronomía. Universidad de la República. Estación Experimental Mario A. Cassinoni (EEMAC). Ruta 3, km 363, 60000. Paysandú, Uruguay. Correo electrónico: toto@fagro.edu.uy.
Recibido: 19/9/11 Aceptado: 27/9/12
Resumen
El trabajo se realizó en la Facultad de Agronomía, ubicada en Paysandú, Uruguay (32º 23’57,86" latitud sur y 58º 02' 42,48" longitud oeste, 61 msnm). El objetivo fue estudiar la respuesta en la producción invernal de un campo natural bajo diferentes dosis de fertilización nitrogenada y ofertas de forraje. El experimento fue conducido en un diseño experimental central compuesto rotacional de precisión uniforme, con dos factores a cinco niveles cada uno: nitrógeno (N) = 0, 44, 150, 256, 300 kg/ha de N y ofertas de forraje (OF) = 4,0; 5,5; 9,0; 12,5 y 14,0 kg MS/100 de peso vivo (PV), distribuidos en dos bloques. Los resultados obtenidos permiten establecer la clara interacción entre el efecto de la fertilización nitrogenada, la intensidad del pastoreo y la composición botánica de la pastura, determinando resultados distintos según los niveles de cada factor utilizado. Existió una marcada repuesta del número de macollos de Bromus auleticus al aumentar la OF de forraje independientemente del nivel de N, que sugiere su baja tolerancia al aumento en la intensidad de pastoreo. Se destaca la capacidad pionera de colonizar ambientes pobres que presentó Stipa setigera, que permitiría la recuperación temprana de ambientes altamente disturbados, posibilitando el incremento de la productividad del sistema.
Palabras clave: DENSIDAD DE PLANTAS, MACOLLAJE, FERTILIZACIÓN, OFERTA DE FORRAJE
Summary
Winter Population Response of Two Native Grasses to Nitrogen Fertilization and Herbage Allowance
The research was conducted at the School of Agronomy, located in Paysandú, Uruguay (32º 23’57,86" South and 58º 02' 42,48" West 61 msnm). The aim was to study the response in winter production of a native pasture to different levels of nitrogen fertilization and herbage allowance. The experiment was led in an experimental central compound rotational design of uniform precision, with two factors with five levels each: nitrogen (N) (0, 44, 150, 256, 300 kg/ha of N) and herbage allowance (HA) (4,0; 5,5; 9,0; 12,5 and 14,0 kg MS/100 kg of live weight LW), distributed in two blocks. The results obtained show a clear interaction between the nitrogen fertilization, grazing intensity and pasture botanical composition, determining distinct results depending on the levels of each factor used. There was a strong response in the number of tillers of Bromus auleticus when increasing the HA regardless of the level of N, suggesting their low tolerance to increased grazing intensity. It highlights the pioneering capacity to colonize poor environments Stipa setigera presented, that would allow early recovery of highly disturbed environments, enabling increased system productivity.
Key words: NUMBER OF PLANTS, TILLERING, FERTILIZATION, HERBAGE ALLOWANCE
Introducción
La vegetación del Uruguay se puede considerar predominantemente herbácea, con una variedad de especies que cambian su frecuencia y sus hábitos morfofisiológicos según las condiciones geológicas, edáficas, topográficas y de manejo a las cuales son sometidas (Berreta, 1996). En el contexto de esta gran diversidad florística existen características comunes a las pasturas naturales: a) baja proporción de leguminosas y b) relaciones de gramíneas invernales y estivales que favorecen generalmente a las segundas (Carámbula, 1997). Por otra parte, c) los suelos en los cuales se apoyan son moderadamente ácidos y poseen contenidos bajos de nutrientes, limitando seriamente su productividad (Zamalvide, 1998). Esas tres características determinan una escasa producción invernal, en promedio 380 kg MS/ha (Boggiano et al., 2005), y baja calidad del forraje ofrecido en primavera-verano (Carámbula, 1997). Al mismo tiempo, el pastoreo con una carga constante de ovinos, vacunos y equinos, ha provocado la degradación y la consecuente disminución de la productividad de las pasturas naturales fundamentalmente en invierno (Carvalho et al., 2011). Esta situación determina que el punto de partida para mejorar la productividad del ecosistema pastoril pase indefectiblemente por un ajuste de la carga animal, restringida por la disponibilidad de forraje otoño-invernal (Millot, 1991).
El objetivo general del presente trabajo fue evaluar la respuesta poblacional otoño invernal, cuantificada a través del número de plantas y macollas, de dos gramíneas nativas de excelente valor forrajero, Stipa setigera Presl. = Stipa neesiana Trin & Rupr. y Bromus auleticus Trin., bajo distintas dosis de fertilizante nitrogenado y ofertas de forraje, dado que ambos factores son de los principales condicionantes de la producción pastoril.
Materiales y métodos
Descripción del sitio experimental
El experimento fue realizado en el invierno del 2003, en la Estación Experimental Mario A. Cassinoni (Facultad de Agronomía), en el km 363 de la ruta 3 General Artigas, departamento de Paysandú, Uruguay (32º 23’57,86" latitud sur y 58º 02' 42,48" longitud oeste, 61 msnm). Los suelos del área experimental son Brunosoles Subeutricos típicos, encontrándose Solonetz como suelos asociados (Altamirano et al., 1976), pertenecientes a la unidad San Manuel de la Clasificación de Suelos del Uruguay. El relieve es de pendientes moderadas y lomadas suaves (Durán, 1985). El análisis de suelo realizado a principios de marzo de 2003 indicó un ph: 5,8, M.O: 5,2 %, P2O5: 6,1 ppm y 5,5 ppm de nitratos.
El área está destinada a la cría de ganado desde hace más de 20 años, manteniéndose como «campo virgen», según sugiere la presencia de varias especies indicadoras que así lo caracterizan: Bromus auleticus, Dorstenia brasiliensis y Geranium albicans (Rosengurtt, 1979). La vegetación presenta un estrato alto dominado por especies arbustivas características, siendo Acacia caven (Espinillo) la dominante. Conjuntamente con estos arbustos, aparece un estrato bajo herbáceo pastoreado dominado por gramíneas de variable valor pastoril que incluyen las especies evaluadas más otras estivales donde se destacan las del género Paspalum.
Diseño experimental
Se estudió el efecto de la oferta de forraje y de la fertilización nitrogenada sobre: el número de plantas y macollos de Stipa setigera y Bromus auleticus.
Para llevar adelante este estudio se aplicaron cinco ofertas de forraje (OF): 4,0; 5,5; 9,0; 12,5 y 14,0 kg MS/ 100 kg de peso vivo animal por día y cinco niveles de fertilización nitrogenada (N) 0; 44; 150; 256; 300 kg de nitrógeno por ha/año.
El rango de ofertas evaluado se fijó en base a resultados de experimentos de la región que determinan los extremos en donde la pastura puede ser degradada por sobre y subpastoreo (Maraschin, 2001) y los niveles intermedios resultan de la aplicación del delineamiento experimental.
Como nivel mínimo de nitrógeno o testigo se consideró la situación normal de producción en la cual no se realiza incorporación. Como nivel superior se consideró un valor máximo que permitiera expresar el potencial otoño-invernal de producción, dada la presencia de especies nativas de alto valor forrajero, como por ejemplo Bromus auleticus, Stipa setigera, Paspaplum dilatatum y Paspalum notatum; los niveles 44; 150; 256; son consecuencia del delineamiento experimental. Las fertilizaciones fueron realizadas al voleo en cuatro aplicaciones distribuidas en otoño-invierno, la primera de ellas el 7 de marzo, la segunda el 22 de abril, mientras que la tercera y cuarta se hicieron el 6 de junio y 21 de julio respectivamente.
El diseño experimental utilizado fue el central compuesto (Cochran y Cox, 1957), con Precisión Uniforme, Rotacional y con los tratamientos distribuidos en dos bloques, en forma ortogonal, de manera que las estimativas de los parámetros del modelo de superficie de respuesta no fueran afectadas por el efecto de los bloques. Para realizar el bloqueo correspondiente al diseño experimental se caracterizó al inicio del experimento la composición botánica mediante el estudio fitosociológico a través del método de punto-transecta (Daget y Poissonet, 1971). Estos datos fueron procesados en planillas en las que se establecieron las frecuencias relativas y absolutas de las especies, para posteriormente ser analizadas estadísticamente. El análisis estadístico se realizó mediante taxonomía numérica: el coeficiente de asociación de Jaccard, con aplicación de análisis de Cluster para agrupar pasturas con mayor similitud taxonómica. Mediante este análisis se agruparon las parcelas en dos bloques diferenciados por su proporción de Bromus auleticus y Stipa setigera, en más y menos de 35%. Los tamaños de los potreros variaron desde 1038 hasta 2219 m2 y el área total fue de 2,13 ha.
El proceso de análisis de los datos se inició con el estudio de la distribución normal de los residuos según el test Shapiro-Wilks. Todas las variables estudiadas satisficieron la condición de normalidad. Para analizar los efectos de ambos factores sobre cada variable se utilizó análisis de varianza. Para estimar las respuestas de cada variable a la fertilización y la asignación de forraje se estudió la superficie de respuesta.
Manejo del experimento y mediciones sobre la pastura
El período evaluado comprende el otoño-invierno del 2003 (1 de marzo al 31 de agosto). Las dosis de nitrógeno fueron aplicadas en su totalidad, en forma de urea, además se corrigió con aplicaciones al voleo de 100 kg/ha de superconcentrado (0-40/41-0+3S) junto con la primer aplicación de urea para alcanzar 10 ppm de P2O5 en el suelo.
Los pastoreos se realizaron con novillos Holstein; los ciclos de pastoreos fueron establecidos en base a resultados experimentales de manejo de pasturas naturales similares (Boggiano et al., 2005), utilizándose en el período experimental ciclos de 50 días con períodos de pastoreo de cinco días y períodos de descanso de 45 días, dependiendo de la oferta de forraje disponible en el momento de ingresar los animales. El manejo durante el verano previo al inicio del período evaluado fue realizado con ciclos de 40 días de pastoreo con períodos de descanso de 35 días, mientras que en la primavera previa fue con ciclos de 35 días de pastoreo y 30 de descanso.
Para cuantificar el número de plantas y macollos de Bromus auleticus y Stipa setigera y comparar su respuesta en cada tratamiento a la dosis de fertilizante nitrogenado y oferta de forraje, se utilizó un círculo de 0,2 m de diámetro, realizándose 15 determinaciones por transecta en tres transectas fijas por parcela a finales de julio del 2003.
Resultados
Número de plantas de Bromus auleticus
La población otoño-invernal de plantas/ m2 ajustó significativamente al modelo de superficie de repuesta: y = 99,5 +0,6 N –20,1 OF –1,8x10-3 N2 + 6,0x10-3 OFxN +1,3 OF2; (R2 = 0,86). Se detectaron efectos significativos lineales y cuadráticos para la oferta de forraje y nitrógeno (Cuadro 1).
La población otoñal de plantas de Bromus auleticus aumentó desde OF medias hacia valores extremos, mientras N presentó valores máximos en las dosis medias, independiente del nivel de OF. El valor máximo de 125 plantas/m2 se logró con 164 kg N/ ha y 14 % de OF (Figura 1).
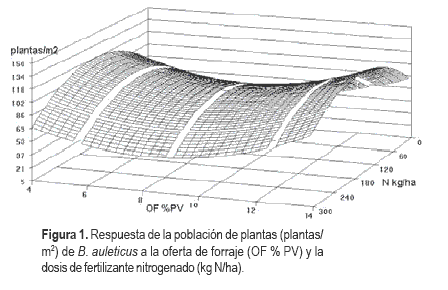
Número de macollos de Bromus auleticus
La población de macollos/m2 ajustó al modelo de superficie de respuesta y = -123,0 + 8,3 N - 30,7 OF - 0,02 N2 - 0,3 OFxN +10,7OF2; (R2 = 0,96). Se detectaron efectos significativos cuadráticos para la oferta de forraje y la dosis de nitrógeno (Cuadro 1).
Si bien la respuesta al agregado de N fue significativa, la misma fue de baja magnitud indicando que las máximas poblaciones de macollos podrían lograrse con aplicaciones medias. Por otro lado se encontró una marcada respuesta al aumento en el número de plantas a la reducción en la intensidad del pastoreo, alcanzando poblaciones de 1750 macollos /m2 con OF de 14 %PV y 132 kg/ha de N (Figura 2).
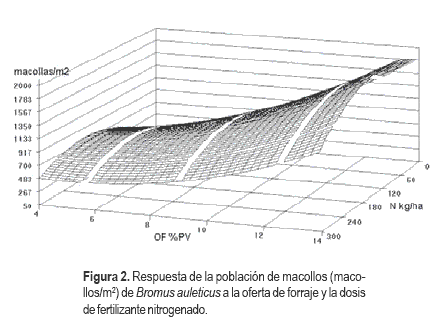
Número de plantas Stipa setigera
La densidad otoño-invernal de plantas (plantas/m2) ajustó al modelo de superficie de respuesta: y = 229,97 – 9,84 OF - 0,94 N - 0,05 OF2 + 0,04 NxOF + 0,001 N2; (R2 = 0,84); se detectaron efectos significativos lineal y cuadrático para la dosis de nitrógeno y la interacción NxOF (Cuadro 1). El valor máximo de densidad de plantas estimada fue de 162 plantas/m2, con OF= 6,5 % PV y N= 3,3 kg N/ha (Figura 3).

El efecto de la interacción fertilizante y de la intensidad del pastoreo se observa en la variación de la densidad a medida que aumenta la oferta de forraje a dosis variables de nitrógeno. A niveles altos de N (300 kg/ha) el número de plantas disminuye cuando aumenta la OF, mientras que a niveles bajos de N (0 kg/ha) el número de plantas también disminuye a niveles altos de OF (Figura 3).
Número de macollos de Stipa setigera
El número de macollos/m2 ajustó significativamente al modelo de superficie de respuesta y = -1163,5 + 379,5 OF + 16,4 N – 15,6 OF2 –1,0 NxOF – 2*10-2 N2 (R2 = 0,78). Se detectaron efectos significativos lineales y cuadráticos de la oferta de forraje y dosis de N y para la interacción N x OF (Cuadro 1).
La interacción OFxN indica que con altas ofertas de forraje el incremento en las dosis de nitrógeno produjo una reducción del macollaje, mientras que lo opuesto ocurrió con bajas ofertas de forraje. Asimismo con bajas dosis de N el macollaje aumentó a medida que aumentó la oferta de forraje, mientras que lo opuesto ocurrió con altas dosis de fertilizante. El máximo número de macollos (1800 macollos/m2) se registró a mínimos de OF (4,2 % PV) y altas dosis de nitrógeno (260 kg N/ha). Es de destacar que en el extremo opuesto, alta oferta y baja dosis de fertilizante, la densidad de macollos de Stipa setigera fue también alta (Figura 4).
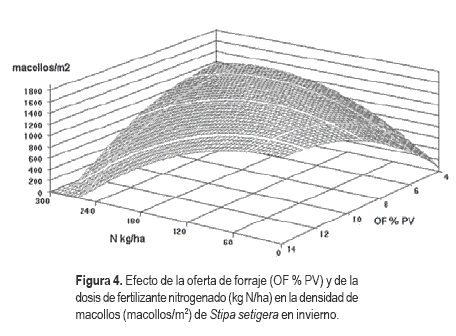
Discusión
El efecto de las ofertas de forraje con respecto al número de plantas de Bromus auleticus, fue consecuencia de la combinación de dos efectos: por un lado la capacidad de emergencia y rebrote otoñal, y por otro su capacidad de sobrevivencia en el período estival. En este sentido los manejos más intensos (OF menores) durante el verano redujeron más la competencia de las estivales ya que se encontraban en período de crecimiento (Carvalho et al., 2008), mientras que Bromus auleticus fue menos perjudicado por presentar semi-latencia estival (Rosengurtt, 1946). Esto determinó que en otoño las parcelas de menor oferta presentaran un tapiz estival menos competitivo para el crecimiento inicial de las invernales, lo que concuerda con lo expresado por Gallinal et al. (1938). En contraposición, en las condiciones de altas ofertas de forraje se promovieron las cespitosas de porte alto como Bromus auleticus, que mantuvieron sombreadas a las estivales desde la primavera anterior reduciendo su cobertura (Boggiano y Zanoniani, 2003).
En efecto, las estivales permanecen bajas durante el verano por efecto del sombreado de los restos secos que se acumulan y son poco removidos por el pastoreo, ya que los animales rechazan el material muerto, que permanece hasta el otoño. Se debe considerar que la especie estival de mayor cobertura en esta área es Paspalum notatum, que no es buena competidora en condiciones de sombreado. Esto concuerda con lo expresado por Altesor et al. (2006) y Formoso y Colucci, (2003), quienes encontraron una mayor proporción de Stipa setigera y otras gramíneas cespitosas y/o erectas en parcelas no pastoreadas, frente a la dominancia de gramíneas postradas estivales como Paspalum notatum, Axonopus affinis y Stenotaphrum secundatum y malezas enanas, con escasa presencia de gramíneas tiernas-finas en las parcelas pastoreadas.
De esta forma se determina que se llegue al otoño con mayor población de plantas de Bromus auleticus en las ofertas de forraje (OF) mayores. En OF medias se tendría una condición más favorable para la competencia de las estivales, determinado poblaciones de Bromus auleticus menores. La marcada respuesta del número de macollos de esta especie al aumentar la OF de forraje independientemente del nivel de N, sugiere una baja tolerancia de la misma al aumento en la intensidad de pastoreo y su menor probabilidad de encontrarse en los campos naturales del Uruguay. Esto concuerda con lo expresado por Lemaire y Chapman (1996), Eggers et al. (2004) y Gastal et al. (2004), quienes expresan que con pastoreo el equilibrio entre aparición y muerte de macollos es fuertemente dependiente del régimen de defoliación, que determina el índice de área foliar (IAF), que influencia las interacciones competitivas por luz y por nutrientes y determinante de su capacidad de sobrevivencia. De esta forma al ser Bromus auleticus una planta cespitosa erecta, los pastoreos muy severos reducirían las reservas de carbohidratos ubicadas en las vainas de las hojas (Richards, 1993), provocando su desaparición de las pasturas naturales (Millot, 1991).
En Stipa setigera se encontró una mayor densidad de plantas en los tratamientos con bajos niveles de N y de OF, lo que podría responder a una moderada capacidad de competencia inicial en ambientes intensamente pastoreados siempre y cuando existan bajos niveles de fertilidad. En ambientes más pobres encontraría una vegetación menos densa y menos competitiva, permitiendo la instalación de nuevas plantas, favorecida por su sistema de dispersión por sus frutos, lo que fomentaría la idea de su capacidad de colonizadora temprana en ambientes altamente disturbados. Al aumentar la fertilidad del suelo y manejar altas intensidades pierde capacidad de competencia frente a otras especies de postradas como Paspalum notatum, que domina estas áreas y conforma una cobertura densa que dificulta la instalación de nuevas plantas. Se debe tener en cuenta que para esta especie se han logrado respuestas al fertilizante hasta 250 kg/ha N y producción de forraje superior a los 14000 kg MS/ha, lo que permite dar una idea de su capacidad de competencia (Boggiano, 2000; Boggiano et al., 2001; Steiner, 2006). En condiciones opuestas (alta oferta y alto nivel de nitrógeno) la densidad de plantas aumenta, dado que el nutriente no es limitante y esta especie de hojas finas y macollos erectos puede instalarse mejor, dada la mayor luminosidad que llega al estrato inferior al estar el tapiz más abierto, y de competir mejor con Paspalum notatum de hábito postrado. Cuando existen bajos niveles de nitrógeno la población disminuye al aumentar la OF debido a que encuentra la competencia de otras especies cespitosas que se comportan mejor en estos ambientes menos iluminados, como ya fue mencionado para el caso de Bromus auleticus.
El número de macollos de Stipa setigera aumentó a altos niveles de N al incrementarse la intensidad de pastoreo, resultando la mayor fertilidad y el ambiente lumínico de mayor calidad de radiación en plantas más macolladas. En cambio a altas asignaciones de forraje en ambientes más fértiles se produjo un exceso de forraje que generó sombreado y por lo tanto una baja relación R/ RL (Rojo/Rojo lejano) en la base de los macollos (Deregibus et al., 1985). En estas condiciones el macollaje se redujo, indicando que la planta está sufriendo cierto grado de competencia por luz debiendo priorizar el crecimiento en altura en detrimento del macollaje, coincidiendo con lo mencionado por Robson (1967). Por otro lado, a bajos niveles de N, los macollos aumentaron al reducir la intensidad de pastoreo (mayores OF), como consecuencia de mantener mayor área foliar y mayor disponibilidad de N dentro de la planta, que puede ser redistribuido para formar nuevos macollos posteriormente al pastoreo (Briske y Richards, 1995). Con una menor disponibilidad de nitrógeno, la reposición del área fotosintéticamente activa fue más lenta, por lo que es necesaria una mayor área foliar remanente para mantener similar ritmo de acumulación de forraje en el proceso de rebrote, lo cual coincide en parte con los trabajos de Chapman et al. (1983), quienes encontraron mayor densidad de macollos de Lolium perenne cuando fue pastoreado con carga rotativa que con carga continua. Los resultados obtenidos son similares a los mencionados por Chapman y Lemaire (1993), Lemaire y Chapman (1996) y por Eggers et al. (2004) en cuanto a la regulación que se puede lograr con pastoreo, ya que el equilibrio entre aparición y muerte de macollos es fuertemente dependiente del régimen de defoliación, determinando el índice de área foliar (IAF) que influencia las interacciones competitivas por luz y por nutrientes.
Finalmente, los resultados obtenidos permiten establecer la clara interacción entre el efecto que posee la fertilización nitrogenada, la intensidad del pastoreo y la composición botánica de la pastura, obteniéndose resultados distintos según los niveles de cada factor utilizado. Estos resultados también determinan que el número de plantas y macollos sea resultado del balance entre las tasas de aparición y muerte de macollos (Boggiano, 2000; Quadros et al., 2006) como consecuencia del efecto del pastoreo sobre la heterogeneidad de la pastura (Bullock y Marriot, 2000; Rook y Tallowin, 2003), de la composición florística y del momento de aplicación del fertilizante (Chapin, 1980; Tilman, 1988; Berendse et al., 1992). Como consecuencia de ello se debe relativizar el uso del nitrógeno sin previamente conocer la composición botánica de la pastura y el manejo ganadero utilizado (Boggiano y Zanoniani, 2003; Van der Hoek, 2011).
Conclusiones
Los resultados obtenidos marcan interacciones claras entre la intensidad de pastoreo, la fertilización nitrogenada y la composición de la pastura. Sin embargo, existieron dos aspectos sumamente importantes a destacar: por un lado la marcada repuesta del número de macollos de Bromus auleticus al aumentar la OF de forraje independientemente del nivel de N, que sugiere su baja tolerancia al aumento en la intensidad de pastoreo, y por otro lado, la capacidad pionera de colonizar ambientes pobres que mostró Stipa setigera, lo cual permitiría la recuperación temprana de ambientes altamente disturbados, posibilitando el incremento de la productividad del sistema.
Bibliografía
Altamirano A, da Silva H, Durán A, Echevarría A, Panario D, Puentes R. 1976. Carta de reconocimiento de suelos del Uruguay : Clasificación de suelos del Uruguay. Montevideo : MAP. 96p.
Altesor A, Piñeiro GF, Lezama F, Jackson RB, Sarasola M, Paruelo JM. 2006. Ecosystem changes associated with grazing in subhumid South American grassland. Journal of Vegetation Science, 17: 323 – 332.
Berendse F, Elberse W, Geerte RH. 1992. Competition and nitrogen loss from plants in grassland ecosystems. Ecology, 73: 46 – 53.
Berreta E. 1996. Campo natural : Valor nutritivo y manejo. En: Producción y manejo de pasturas. Montevideo: INIA. (Serie Técnica ; 80). pp. 113 – 128.
Boggiano P. 2000. Dinâmica de produção primaria da pastagem nativa em área de fertilidade corrigida sob efeito de adubação nitrogenada e oferta de forragem [Tese Doutorado]. Porto Alegre : Facultade de Agronomía, Universidade Federal do Río Grande do Sul. 191p.
Boggiano P, zanoniani R, millot JC. 2005. Respuestas del Campo Natural a niveles crecientes de intervención. En: Seminario de actualización técnico en manejo de campo natural. Montevideo: INIA. (Serie Técnica; 151). pp. 105 – 113.
Boggiano P, Zanoniani R. 2003. Alternativas para promover las especies deprimidas del Campo Natural: Respuesta otoñal en la estructura poblacional de Bromus auleticus Trinus AL. En: I Encuentro de Ecología del Uruguay. Montevideo : Sociedad Zoológica del Uruguay. p. 107.
Boggiano P, Maraschin GE, Nabinger C. 2001. Herbage allowance and nitrogen fertilization effects on morphological characteristics of Paspalum notatum Flügge. En: 19th International Grassland Congress; 11 - 21 febrero 2001; São Pedro, São Paulo, Brazil. Piracicaba : FEALQ. pp. 60 – 61.
Briske D, Richards J. 1995. Plant responses to defoliation: a physiological, morphological and demographic evaluation. En: Bedunah D, Sosebee R [Eds.]. Wildland plants : physiological ecology and developmental morphology. Denver : Society of range management. pp. 635 – 710.
Bullock JM, Marriot CA. 2000. Plant responses to grazing and opportunities for manipulation. En: Rook AJ, Penning PD. [Eds.]. Grazing management, the principles and practice of grazing, for profit and environmental gain, within temperate grassland systems. Reading : British Grassland Society. pp. 17 – 26.
Carámbula M. 1997. Pasturas naturales mejoradas. Montevideo : Hemisferio Sur. 524p.
Carvalho PCF, Nabinger C, Lemaire G, Genro TCM. 2011. Challenges and opportunities for livestock production in natural pastures : the case of Brazillian Pampa Biome. En: Feldman SR, Oliva GE, Sacido MB [Eds.]. IX International Rangeland Congress; 2011; Rosario, Argentina. Rosario: Universidad de Rosario. pp. 9 – 15.
Carvalho PCF, Mezzalira JC, Fonseca L. 2008. Características estruturais do pasto e o consumo de forragem: o quê pastar, quanto pastar e como se mover para encontrar o pasto. En: Pereira OG, Obeid JA, Fonseca DM, Nascimento JRD [Eds ]. IV Symposium on strategic management of pasture, and II International symposium on animal production under grazing; Viçosa, Anais. Viçosa: Universidad Federal de Viçosa. pp. 101 – 130.
Chapin ES. 1980. The mineral nutrition of wild plants. Annual Review of Ecology and Systematics, 11: 233 – 260.
Chapman DF, Lemaire G. 1993. Morphogenetic and structural determinants of plant regrowth after defoliation. En: 17th International Grassland Congress; 1993. Palmerston North : New Zeland Grassland Association, Keeling & Mundi. pp. 95 – 104.
Chapman DF, Clark DA, Land CA, Dymock N. 1983. Leaf and tiller growth of Lolium perenne and Agrostis spp. and leaf appearance rates of Trifolium repens in set-stocked and rotationally grazed hill pastures. New Zeland Journal of Agricultural Research, 26: 159 – 168.
Cochran WG, Cox MG. 1957. Experimental designs. 2nd ed. New Cork: John Wiley. 611p.
Daget Ph, Poissonet J. 1971. Une méthode d´analyse phytologique de prairies: Critéres d´application. Annales Agronomiques, 22(1): 5 – 41.
Deregibus VA, Sanchez RA, Casal JJ, Trlica MJ. 1985. Tillering responses to responses to enrichment of red light beneath the canopying a humid natural grassland. Journal of applied Ecology, 22: 199 – 206.
Duran A. 1985. El medio ambiente. En: Los suelos del Uruguay. Montevideo : Hemisferio Sur. pp. 47 – 64.
Eggers L, Cadenazzi M, Boldrini II. 2004. Phyllochron of Paspalum notatum FL. and Coelorhachis selloana (HACK.) Camus in Natural Pasture. Scientia Agricola, 61(4): 353 – 357.
Formoso D, Colucci P. 2003. Vegetation changes in native plant comunities in basaltics deep an shallow soils after 10 years of grazing exclosure in Uruguay. En: VII International Rangeland Congress. Durban, South Africa. Durban: Rangeland Congress. pp. 327 – 330.
Gallinal H, Bergalli L, Campal E, Aragone L, Rosengurtt B . 1938. Estudios sobre praderas naturales del Uruguay : Primera Contribución. Montevideo : Imprenta Germano Uruguaya. 208p.
Gastal F, Lemaire G, Lestienne F. 2004. Defoliation, shoot plasticity, sward structure and herbage utilization [CD ROM]. En: II Simposio em Ecofisiologia das Pastagem e Ecologia do Pastejo. Curutiba: Universidad de Curitiba.
Lemaire G, Chapman D. 1996. Tissue flows in grazed plant communities. En: Hodson J, Illius AW. [Eds.]. The ecology and management of grazing systems. Wallingford: CAB International. pp. 3 – 36.
Maraschin GE. 2001. Production potential of South América grasslands. En: 19th International Grassland Congress; 11 - 21 febrero 2001; São Pedro, São Paulo, Brazil. Piracicaba: FEALQ. pp. 5 – 15.
Millot JC. 1991. Manejo del Pastoreo y su incidencia sobre la composición botánica y productividad del Campo Natural. En: Pasturas y producción animal en áreas de ganadería extensiva. Montevideo: INIA. (Serie Técnica; 13). pp. 68 – 70.
Quadros FLF, Cruz P, Theu JP, 2006. Uso de tipos funcionais de gramíneas como alternativa de diagnóstico da dinâmica e do manejo de campos naturais [CD ROM]. En: 43 Reunião anual da sociedade brasileira de zootecnia; João Pessoa. João Pessoa : SBZ.
Richards JH. 1993. Physiology of plants recovering from defoliation. En: Proceedings of the XVII International Grasslands Congress. Palmerston North New Zeland: Keeling & Mundey. pp. 85 – 93.
Robson M. 1967. A comparision of British and North American varieties of tall fescue. Journal Applied Ecology, 4: 475 – 484.
Rook AJ, Tallowin JRB. 2003. Grazing pasture management for biodiversity benefit. Animal Research, 52: 181-189.
Rosengurtt B. 1979. Tablas de comportamiento de las especies de plantas de Campos Naturales en el Uruguay. Montevideo: Universidad de la República. 86p.
Rosengurtt B. 1946. Estudios sobre Praderas Naturales del Uruguay: 5.ª Contribución. Montevideo : Rosgal. P 249.
Steiner MG. 2006. Caracterização agronômica, molecular e morfológica de acessos de Paspalum notatum Flugge e Paspalum guenoarum Arech [Tesis de maestría]. Porto Alegre : Universidade Federal do Rio Grande do Sul. 129p.
Tilman D. 1988. Plant strategies and the structure and dynamics of plants communities. Princeton : Princeton University. 360p.
Van der Hoek KW. 2011. Nitrogen budgest as tool for sustainable rangelands. En: Feldman SR, Oliva GE, Sacido MB. [Eds.]. IX International Rangeland Congress; 2011; Rosario, Argentina. Rosario: Universidad de Rosario. p. 355.
Zamalvide J. 1998. Fertilización de Pasturas. En: Anales XIV reunión del grupo técnico regional del Cono Sur en mejoramiento y utilización de los recursos forrajeros del área tropical y subtropical: Grupo Campos. Montevideo: INIA. (Serie Técnica; 94). pp. 97 - 107.

