










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Links relacionados
Compartilhar

 Permalink
PermalinkAgrociencia (Uruguay)
versão impressa ISSN 1510-0839versão On-line ISSN 2301-1548
Agrociencia Uruguay vol.14 no.1 Montevideo jan. 2010
Intensidad del pastoreo sobre la estructura de una pastura de Lolium perenne cv Horizon
Saldanha, Sylvia1, Boggiano, Pablo2 y Cadenazzi, Mónica2
1 Facultad de Agronomía, Est. Exp. Salto. Correo electrónico: sylsal71@adinet.com.uy
2 Facultad de Agronomía, Est. Exp. Dr. M. A. Cassinoni.
Recibido: 1/9/09 Aceptado: 15/3/10
Resumen
Se evaluó el efecto de intensidades de pastoreo aplicadas como ofertas de forraje (OF) de 2,0 - 4,5 – 7,0 y 9,5 kg MS/100 kg de peso vivo animal, de agosto a noviembre, en la densidad y peso de los macollos de una pastura de Lolium perenne. El diseño fue completamente al azar con dos repeticiones. Las relaciones entre ambas variables y las OF fueron lineales en octubre y noviembre: Densidad de macollos (macollos/m2)=1821+209 OF (P<0,001; r2=0,76), Peso seco del macollo (mg)=19,3+8,1 OF (P<0,0001; r2=0,70), no ocurriendo mecanismos de compensación. Las OF afectaron la persistencia de la pastura, perdiéndose en noviembre 58 plantas/m2 por cada 1 % de disminución en la OF (P<0,0001; r2=0,84). Las OF también afectaron el rebrote. La relación entre el peso del macollo remanente y el del rebrote para todo el período fue significativa (P<0,0004; r2=0,66): Peso de macollo (mg) pre-pastoreo=34,61+0,99 x, siendo x=peso de macollo del forraje remanente del pastoreo anterior. Los resultados obtenidos reflejan la complejidad de los procesos, y que en las condiciones de Uruguay la luz no es el principal factor limitante en la productividad de las pasturas, y que la intensidad del pastoreo es una variable importante de manejo.
Palabras clave: raigrás perenne, oferta de forraje, peso macollos, densidad macollos
Summary
Effect of the intensity of defoliation in the structure of pasture of Lolium perenne cv Horizon
The effect of intensities of grazing applied as forage offers (OF) - 2,0 - 4,5 - 7,0 and 9,5 kg DM/100 kg of animal liveweight on tillers density and dry weight was evaluated from August to November in a Lolium perenne sward. The design was totally at random with two replicates. The relationships between both variables and the OF were lineal in October and November: Tillers density (tillers/m2)=1821+209 OF (P<0,001; r2=0,76), Tillers dry weight (mg)=19,3+8,1 OF (P<0,0001; r2=0,70). No compensatory mechanisms occurred. Grazing intensity affected the persistence of the pasture. In November, 58 plants/m2 got lost for every 1 % decrease in the OF (P<0,0001; r2=0,84). The OF also affected pasture regrowth. The relationship between tiller weight of the remainder pos-graze and tiller weight of the regrowth for the whole period was significant (P<0,0004; r2=0,66): Tillers dry weight (mg) pre-graze=34,61+0,99 x, for x=weight of tiller of the forage remainder of the previous defoliation. The obtained results reflect the complexity of the processes. Under Uruguay´s conditions the light is not the main factor limiting pasture productivity, while defoliation intensity is an important variable.
Key words: Lolium perenne, forage offer, tiller weight, tiller density
Introducción
El manejo de las pasturas, dieta básica de vacunos y ovinos en el Uruguay, es primordial para mantener una alta productividad y calidad de forraje en el tiempo.
Ante variaciones en el manejo y/o en las condiciones ambientales, las pasturas responden modificando su estructura a través de procesos fisiológicos. La estructura de la pastura determina la productividad primaria y secundaria de los ecosistemas pastoriles (Gastal et al., 2004), pues afecta su crecimiento (al determinar el IAF), su persistencia (a través de la población de macollos) y su utilización por el animal. Las características estructurales en respuesta al ambiente y/o manejos del pastoreo sufren modificaciones progresivas y reversibles (plasticidad fenotípica) con el objetivo de restablecer el equilibrio entre demanda y suministro de recursos (Lemaire y Chapman, 1996). Así la defoliación al remover el área foliar modifica la cantidad y calidad de la luz (aumenta la relación rojo/rojo lejano) que llega a la base de la pastura, respondiendo esta con un aumento en la densidad de macollos (Gautier et al., 1999).
La producción de forraje en gramíneas se puede descomponer en el número de macollos por unidad de área de suelo y el rendimiento individual de cada uno, sin embargo la densidad y tamaño de macollo están negativamente correlacionados (Grant et al., 1983). Se ha observado en diferentes especies una relación de compensación entre tamaño y densidad de macollos en pasturas no defoliadas, que presenta una relación funcional con pendiente de -3/2 a escala logarítmica y se asume que esta se cumple bajo pastoreo (Bircham y Hodgson, 1983).
Estos cambios en la relación tamaño/densidad de macollo, así como la asociación entre tasa de crecimiento y tasa de senescencia foliar, explican los rendimientos constantes de Lolium perenne bajo pastoreo continuo y condiciones ambientales no limitantes (Bircham y Hodgson 1983, Matthew et al., 2000; Gastal et al., 2004).
Bajo condiciones limitantes y/o cambiantes, se modifican las relaciones entre las variables o las tasas en que ocurren los cambios (Agnusdei y Assuero, 2004). En pastoreos intermitentes la variación en el IAF en la pastura es máxima (Lemaire, 2001) lo que determina que el proceso de adaptación sea continuo al variar permanentemente el microambiente lumínico (Grant et al., 1981).
El objetivo de este trabajo fue estudiar, sobre una pastura de Lolium perenne cv. Horizon, las respuestas en el número y peso de los macollos a intensidades de pastoreo aplicadas como OF de 2,0- 4,5 -7,0 y 9,5 kg MS/100 kg de peso vivo animal, bajo un sistema de pastoreo intermitente de agosto a noviembre.
Materiales y métodos
El área experimental se localizó en la Estación Experimental Mario A. Cassinoni de la Facultad de Agronomía de la Universidad de la República (32º 20’S de latitud; 58º 20’W de longitud, a 61m sobre el nivel del mar), Paysandú. El clima en esta zona del Uruguay según Santibáñez (1994) es templado supratermal estenotérmico y regular perhúmedo.
La pastura utilizada fue una mezcla de Lolium perenne cv Horizon, Trifolium repens cv Zapicán y Lotus corniculatus cv San Gabriel sembrada en directa el 30 de abril de 2006 con 15, 2 y 8 kg de semilla/ha respectivamente, sobre un Brunosol eútrico típico háplico de la Unidad San Manuel, de acuerdo a la clasificación de suelos del Uruguay (Altamirano et al., 1976). La gramínea se sembró en línea a 17 cm y se fertilizó a la siembra con 100 kg/ha de 18-46-0 y el 30 de mayo con 50 kg/ha de 46-0-0. La presencia de leguminosas y malezas durante el período abarcado por este estudio fue ínfima.
Las condiciones ambientales desde la siembra se resumen en la Figura 1 y Cuadro 1. En la figura 1 se presenta el resultado de un balance hídrico decádico para el período marzo-noviembre 2006 realizado con la metodología de Thornthwaite y Mather (1955) suponiendo una exploración radicular de 30 cm. Se observa que el agua almacenada en el suelo fue disminuyendo desde la aplicación de los tratamientos hasta un mínimo que coincidió con el segundo pastoreo evaluado (22 de octubre), luego del cual las precipitaciones acontecidas permitieron restaurar el contenido de agua en el suelo.
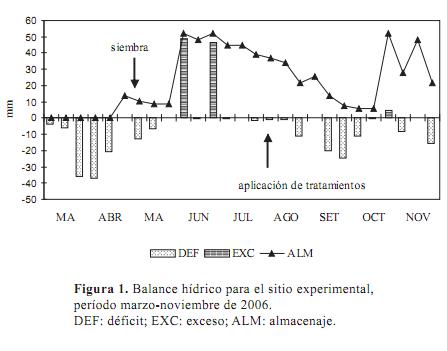
En el Cuadro 1 se presentan las condiciones ambientales entre la siembra y el primer pastoreo y entre éste y los siguientes. En los tres últimos períodos las temperaturas (T) pudieron afectar el crecimiento de Lolium perenne, dado los valores promedio de las T máximas.
Los datos meteorológicos -precipitaciones, T y evaporación del tanque A fueron obtenidos del Aeropuerto Chakling ubicado a menos de 3 km del sitio experimental.
Los tratamientos consistieron en cuatro OF: 2,0; 4,5; 7,0 y 9,5 kg de MS/100 kg del peso vivo animal que se aplicaron el 6 de agosto, 20 de septiembre, 22 de octubre, 10 de noviembre y 30 de noviembre, evaluándose los últimos cuatro pastoreos. Se utilizaron seis novillos tester fijos en cada OF (tres de 250 kg y tres de 350 kg de peso vivo), más volantes para ajustar la misma. El diseño experimental fue completamente al azar con dos repeticiones en cada área, utilizándose ocho parcelas de diferente superficie según tratamientos, para mantener similar densidad animal. Sobre aspectos de la conducción del experimento remitirse a García et al. (2008).
Para el análisis estadístico de los datos fueron utilizadas técnicas de regresión y de análisis de varianza, con medidas repetidas en el tiempo. Se ajustaron modelos de regresión lineales y cuadráticos para cada fecha entre las OF y las variables estudiadas, comparándose las funciones resultantes. En el caso en que las funciones no eran estadísticamente diferentes, se ajustó una función general para todo el período.
En todos los casos se estudió el comportamiento de los errores para los modelos ajustados, mediante técnicas de diagnóstico gráficas y numéricas.
El criterio de significación utilizado en los ajustes de modelos de regresión fue de 10 %. En los casos en que no existió un ajuste significativo de los modelos precedentes, fue realizado un estudio de medias de tratamientos por fechas, considerando medidas repetidas en el tiempo. Este modelo consideró la posible heterogeneidad de varianzas en el tiempo corrigiendo los datos por la matriz de auto correlación temporal UN (SAS 8.1). Las medias de los efectos significativos fueron separadas usando el test de Tukey (P<0,05).
Determinaciones
Muestreo
Previo a cada pastoreo se extraían tres muestras del forraje disponible por parcela. Las muestras consistían en un pan de tierra de 20 cm x 17 cm y de 10 cm de profundidad. Se les extraía la tierra y se lavaban las raíces.
Después de cada pastoreo se medía con regla la altura del forraje rechazado (3 a 5 puntos/muestra) en cinco muestras por parcela de 20 x 17 cm de superficie, luego de lo cual se cosechaba el forraje de cada muestra con tijera de aro al ras del suelo.
Variables
En cada muestra pre pastoreo se contabilizaba el número de plantas y macollos presentes. Los macollos se clasificaban subjetivamente en dos categorías, grandes y pequeños, en función del diámetro de los mismos. Se eliminaban las raíces. Se determinaba el número de macollos de cada conjunto y su masa aérea, colocándose en estufa de aire forzado a menos de 60 ºC hasta peso constante.
En las muestras del forraje remanente al pastoreo se determinaba el número de macollos y su masa, secándolos en estufa hasta peso constante para posteriormente pesarlos.
Resultados y discusión
Densidad de macollos
La densidad de macollos es el producto de la densidad de plantas por el número de macollos por planta. La densidad de macollos promedio en el período fue de 3232 ± 861 macollos/m2.
Las diferencias en densidad se dan muy temprano en el desarrollo y se mantienen después en el tiempo (Skinner y Nelson, 1994). Por lo que la variación en el número de macollos es un proceso lento, principalmente si hay deficiencia de N (Duru y Ducrocq, 2000). En este trabajo los tratamientos se aplicaron a los 98 días pos siembra. La capacidad de adaptación de la estructura de la pastura puede ser dañada notablemente con cambios en el manejo. Por ejemplo: si por estar sin pastoreo mucho tiempo desarrolla una estructura adaptada a sostener una alta biomasa (baja densidad de macollos de gran tamaño), al ser luego pastoreada en forma severa puede limitarse su crecimiento (Parsons y Chapman, 2000).
Luego de dos pastoreos (Figura 2), se encontró una relación funcional de la densidad de macollos con las OF. El ajuste en los tres últimos muestreos fue lineal (P<0,001; r2=0,76) indicando un incremento de 209 macollos/m2 por cada 1% de OF, a partir de 1821 macollos/m2.
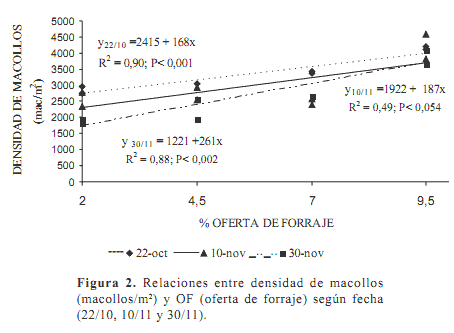
Se observa que con el transcurrir del tiempo fue disminuyendo la densidad de macollos (P<0,0001). La dinámica de los macollos es función de la tasa de aparición y de la tasa de muerte, tasas que difieren con el manejo y la estación (Hernández Garay et al., 1997). La tasa de aparición disminuye a medida que el IAF aumenta (Lemaire y Chapman, 1996) ya que la base de la planta rápidamente es sombreada disminuyendo la relación rojo/rojo lejano. Esta señal se da mucho antes que cualquier reducción en los fotosintatos o de la radiación fotosintéticamente activa interceptada o disponible (Calsal et al., 1990). El pastoreo modifica la cantidad y calidad de la luz que llega a la base de la pastura. A mayor intensidad del pastoreo, aumenta la relación rojo/rojo lejano y las plantas responden con un aumento en la densidad de macollos (Gautier et al., 1999).
Pero si bien la competición por luz es un importante regulador del macollaje, el suministro de C es el factor más importante en la sobrevivencia de los macollos (Bahmani et al., 2000). En Lolium perenne la prioridad en la distribución de los carbohidratos de reserva luego de una defoliación es el rebrote de hojas, luego el reinicio del crecimiento radicular y por último la aparición de nuevos macollos (Donaghy y Fulkerson, 1998).
En este trabajo el estrés impuesto a las plantas en las OF de 2,0 y 4,5 % determinó que la densidad de macollos al 30 de noviembre fuera respectivamente el 47 y 51 % de la densidad al 20 de septiembre, siendo significativo (P<0,0001) el descenso en el número de macollos en el tiempo en estos dos tratamientos.
El macollaje está muy relacionado con los carbohidratos de reserva de las plantas (Fulkerson y Donaghy, 2001). Estos son «movilizados» temprano en la primavera (Parsons y Chapman, 2000), disminuyendo por el rápido crecimiento y alta tasa de respiración de la pastura al aumentar la temperatura (Mathew et al., 2000). La aparición de nuevos macollos se retarda más si los carbohidratos solubles de reserva son bajos (Donaghy y Fulkerson, 1998). Además la muerte de macollos puede ocurrir en jóvenes macollos por inanición de C, incluso antes de emerger (Lemaire y Chapman, 1996).
En las mayores OF hubo mayor reposición (menor competencia por carbohidratos con la producción de láminas) y/o mayor sobrevivencia de macollos, determinando la inexistencia de cambios significativos en el número de macollos.
Según Mathew et al. (2000) en pastoreos con carga intermitente la adaptación morfológica ocurre cuando toda la luz ha sido interceptada, es el final de un proceso de competición por luz. La respuesta lineal detectada en este trabajo puede indicar la falta de competencia por luz, o al menos que no fue este factor la principal causa de competencia.
El estado de desarrollo de los macollos también afecta el balance entre su aparición y muerte. Hernández Garay et al. (1997) observaron que la disminución en la densidad de macollos en Lolium perenne durante la fase reproductiva, fue mayor en la pastura que estuvo cerrada más tiempo.
Si se analizan las variaciones en la densidad de macollos según su tamaño, se observan diferencias en el comportamiento de cada categoría de macollos (grandes y pequeños), si bien ambos aumentaron al aumentar la OF.
Estructura de la población de macollos
Densidad de macollos pequeños
La producción de nuevos macollos es un proceso intermitente que puede ser disparado por la defoliación y la consecuente mejora de las condiciones lumínicas en la base de la pastura. Una menor competencia por carbohidratos entre hojas en desarrollo y macollos, favorece la formación de un mayor número de macollos en los pastoreos más intensos. Esto se manifestó en los primeros pastoreos (Figura 3) en 2,0 y 4,5 % de OF pero estos macollos no sobrevivieron posteriormente, debido también a que el intervalo entre pastoreos fue disminuyendo.
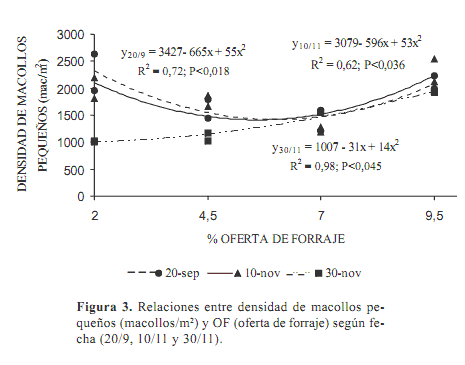
Frente a condiciones poco propicias los macollos de menor tamaño son sacrificados para asegurar la sobrevivencia de los mayores, siendo los más pequeños menos competitivos por los recursos (Hodgson, 1990).
Bircham y Hodgson (1983) determinaron que la tasa de pérdida de macollos es mayor en pasturas con menos de 700 kg MO/ha de biomasa aérea acumulada. En las OF de 2 y 4,5 % el promedio del forraje remanente en el período fue de 320±196 kg MS/ha y 1058±236 kg MS/ha respectivamente. Ong (1978) trabajando con Lolium perenne encontró que las plantas con un alto número de pequeños macollos (menor a 10 mg MS) son las que presentaron pérdida de éstos por muerte, independizándose los macollos totalmente de la importación de asimilatos cuando tienen un peso aproximado de 50 mg MS.
Este proceso se refleja en la evolución del número de macollos pequeños en las dos menores OF (P<0,0014) manifestando una mayor variabilidad en el tiempo.
Densidad de macollos grandes
El número de macollos grandes presentó ajuste lineal con las OF (Figura 4) manifestándose el efecto del pastoreo más tardíamente. La relación fue para el promedio del período, Nº macollos grandes/m2=624+116 OF, (P<0,0001; r2=0,55).
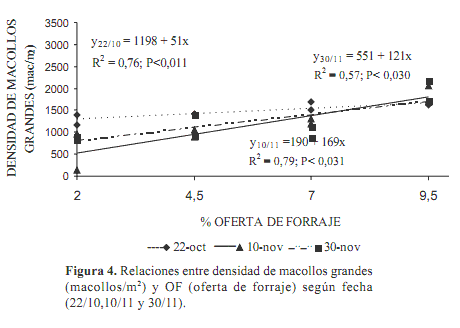
Con alta intensidad de pastoreo el rebrote es más lento y costoso, pues las plantas deben recomponer su aparato foliar con menor cantidad de reservas a partir de menor área foliar remanente y de meristemas menos diferenciados (Briske, 1986) Al ser menores las tasas de crecimiento bajo estos manejos (Fulkerson y Slack 1994, 1995, Lee et al., 2008), menor número de macollos llegan a ser clasificados como «grandes».
El efecto de la OF en la densidad de macollos grandes comenzó a ser significativa (P< 0,017) a principios de noviembre, lo que coincidió con el momento de máximo desarrollo reproductivo. Esta categoría de macollos pesó más en la densidad total de macollos en dicha fecha.
Densidad de plantas
La metodología utilizada para estimar la densidad de plantas puede inducir a imprecisiones en su determinación, pues la planta puede fragmentarse en más de una unidad perdiéndose la identidad original (Parsons y Chapman, 2000).
El número de plantas promedio del período experimental fue de 634 ± 170 plantas/m2. No se detectó efecto de las OF en la densidad de plantas en las dos primeras fechas de evaluación. A partir de noviembre el número de plantas/m2 disminuyó con la intensidad del pastoreo (Nº plantas/m2=356+49 OF; P<0,0006, r2=0,88) y esta reducción se incrementó al pastoreo siguiente (Nº plantas/m2=182+60 OF; P<0,011, r2=0,68). El ajuste (r2=0,84) fue lineal y significativo (P<0,0001) para el promedio de ambos pastoreos, implicando la desaparición de 58 plantas/m2, que representa el 9 % de la densidad promedio, por cada reducción en 1 % en la OF.
Los valores relativos de descenso del número de plantas al final del período experimental con respecto al primer pastoreo (Cuadro 2) fueron similares a la variación relativa en el número de macollos/m2. El crecimiento radicular luego de la defoliación cesa pues la planta prioriza el desarrollo de su capacidad fotosintética. Donaghy y Fulkerson (1998) determinaron que en Lolium perenne el período de cese del crecimiento es más prolongado bajo pastoreos más severos. Una intensidad de corte a 2 cm cada vez que se expande una nueva hoja, provoca que casi la mitad de las raíces no sobrevivan en los primeros 10 días de rebrote. La disminución en el número de plantas en los tratamientos más intensos, pudo deberse a una disminución de la masa radicular (Hernández Garay et al., 2000, Fulkerson y Donaghy 2001; Fulkerson y Slack 1994, Lee et al., 2008) que afecta la sobrevivencia frente a deficiencias hídricas y/o nutritivas del suelo (Fulkerson y Donaghy 2001, Lee et al., 2008). La interdependencia de la raíz y la parte aérea determina que la biomasa radicular en pastoreos más intensos sea menor aumentando la relación parte aérea/raíz con la defoliación. La mayor relación biomasa radicular/biomasa aérea en esta especie fue lograda con pastoreos a 8 cm de altura cuando se evaluó durante 5 meses intensidades de corte de 2 a 16 cm (Hernández Garay et al., 2000).
Número de macollos/planta
En el promedio del período, el número de macollos/planta fue de 5,39 ± 0,97 (Cuadro 3). Es bajo comparado a los obtenidos con dicha especie en invernáculo por Donaghy y Fulkerson (1997), (1998), (30 y 20 macollos/planta respectivamente). En dichos trabajos la densidad de plantas fue menor (96 y 100 plantas/m2 respectivamente) comparada a la obtenida aquí (634 plantas/m2) lo que podría indicar la ocurrencia de mecanismos de compensación (Jewiss, 1972). Pero dada la posibilidad natural de fragmentación de la planta original, que depende del pastoreo y la estación del año (Hume y Brock, 1997) este indicador del proceso de macollaje tiene limitaciones. Para estos autores el promedio de plantas de Lolium perenne en pasturas bajo pastoreo continuo y rotativo fue de 465 plantas/m2, valor más próximo al de este trabajo. Bajo condiciones reales de manejo en el campo Lolium perenne presenta cuatro a cinco macollos promedio/planta según sea manejado con pastoreo continuo o rotativo respectivamente, ya que se fragmentan continuamente las plantas por efecto del pastoreo, con efectos en la supervivencia de las mismas (Brock y Fletcher, 1993).
El número de macollos/planta al final del período (30 de noviembre) fue afectado por los manejos aplicados. Ajustó (r2=0,75) una función lineal (Nº macollos/planta=5,91-0,20 OF) significativa (P<0,011), mostrando un mecanismo de compensación. Altas densidades de plantas presentan menor número de macollos/planta por reducción de la relación rojo/rojo lejano, aún sin existir indicios de competición por luz (Calsal et al., 1990).
Pero el 10 de noviembre la relación (r2=0,63; P<0,048) entre macollos/planta y las OF fue cuadrática (Nº macollos/planta=8,08–1,32 OF+0,105 OF2) con un mínimo de macollos/planta en la OF de 7 %. Esto pudo ser causado por la alta proporción de macollos reproductivos, lo que implica una menor tolerancia al pastoreo (Briske, 1986), manifestándose en una disminución en el número de macollos/planta. La variación en el tamaño de los macollos en una planta es mayor durante la floración, los macollos jóvenes y pequeños reciben menos carbohidratos al modificarse la prioridad de las fosas, siendo pues más sensibles al estrés ambiental ocurriendo la muerte de muchos de ellos (Hodgson, 1990). Esta OF de 7 % presentó siempre una menor proporción de macollos pequeños, indicando menor reposición de macollos. También la alta proporción de macollos reproductivos puede ocasionar una mayor fragmentación de la planta, al desaparecer las conexiones orgánicas entre los macollos (Hume y Brock, 1997).
Peso de macollo
El tamaño de los macollos cambió con la intensidad del pastoreo antes que la variación en la densidad de estos. El peso de un macollo vegetativo depende del número y tamaño de las hojas. El número de hojas formadas determina la tasa potencial de aparición de macollos, siendo la vida promedio de un macollo de raigrás perenne de 100 días, mientras que la tasa de aparición de una hoja es de 13 días (Matthew et al., 2000). Esto determina que los cambios en el número de macollos sean más lentos que el efecto del pastoreo en el crecimiento y senescencia de las hojas (Hepp et al., 1996).
El peso seco de cada macollo fue en el promedio del período de 82,3 ± 43,2 mg.
Al inicio (20 de septiembre) el peso de cada macollo fue significativamente (P<0,0001) mayor que en las siguientes fechas (130,4±43,9 vs 65,5±28,3 mg), ajustando con las OF una función cuadrática (Figura 5). En los tratamientos aliviados la acumulación de forraje era alta y seguramente ya se interceptaba toda la luz incidente (se habría alcanzado el IAF crítico) (Parson y Chapman, 2000). A mayor biomasa la pérdida de carbohidratos por respiración es mayor (Fulkerson y Donaghy 2001), aumenta la tasa de senescencia y la fotosíntesis bruta puede disminuir al reducirse la capacidad fotosintética de las hojas desarrolladas en ambientes más sombríos, lo que determina menores tasas netas de asimilación de C, e incluso un balance negativo de C si el punto de compensación es alcanzado (Parsons y Chapman, 2000). Esto pudo suceder bajo la OF 9,5 %, principalmente por el mayor número de días desde la defoliación anterior (45 días).
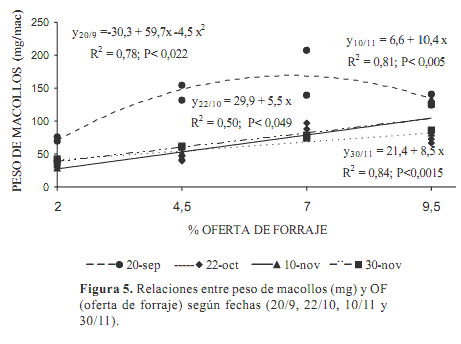
El intervalo entre pastoreos luego del 20 de septiembre disminuyó (32, 19 y 20 días entre pastoreos) y por lo tanto fue menor el forraje acumulado. Al avanzar la primavera la intensidad de luz y el largo del día aumentan, requiriendo mayor área foliar para interceptar el 90 % de la radiación (Parsons y Chapman, 2000). Ambos factores pudieron incidir en que las relaciones entre el peso del macollo y las OF fueran lineales desde el 22 de octubre. Para el conjunto de las tres fechas (22 de octubre y 10 y 30 de noviembre) la regresión fue significativa (P<0,0001; r2=0,70): Peso seco del macollo (mg)=19,3+8,1 OF.
El incremento en el tamaño de macollos al aumentar la OF fue mayor durante el estado reproductivo, lo que en parte estaría asociado a los cambios en la partición de carbohidratos que ocurre en esta etapa: relación parte aérea/raíz y alargamiento de entrenudos (Peacock, 1975).
Peso de macollos grandes
Las relaciones entre el peso de los macollos grandes y las OF fueron similares a las del total de macollos. El peso promedio de esta categoría de macollos fue de 125,6±65,7 mg MS/macollo.
Al inicio por la alta acumulación de forraje la relación fue cuadrática (Figura 6). Los macollos fueron significativamente (P<0,0001) más pesados (196,6±58,0 vs 100,9±48,4 mg MS/macollo) que en los siguientes períodos.
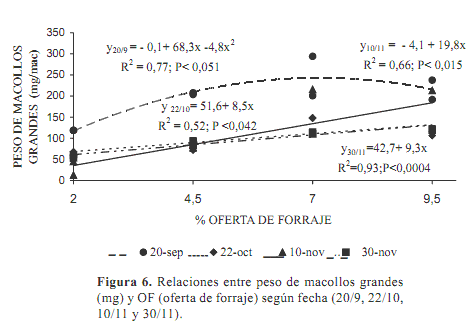
En los pastoreos siguientes la relación fue lineal. La ecuación conjunta para octubre y noviembre fue significativa (P<0,0001) con un r2=0,56 indicando un incremento de 12,9 mg/macollo cada 1 % de OF, a partir de un peso de 28,6 mg.
Peso de macollos pequeños
Los macollos clasificados como pequeños, con un peso promedio de 44,7±19,6 mg MS/macollo, fueron también al inicio (P<0,0009) más pesados (59,7 ± 20,7 mg MS/macollo) que en el resto del período (a excepción del 10 de noviembre). Las relaciones del peso de estos macollos con las OF fueron generalmente lineales (Figura 7).
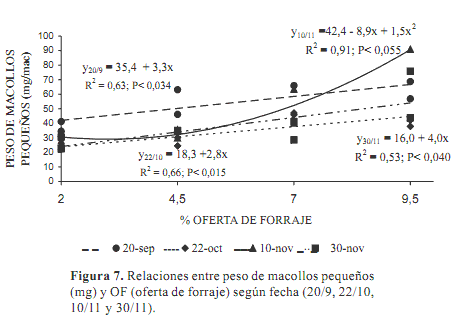
El 10 de noviembre, ya iniciado el período de floración, la relación del peso de los macollos pequeños con el pastoreo fue cuadrática, quizás en las mayores OF muchos de estos macollos estaban alargando los entrenudos y cambiando su estado fenológico.
Relación peso/densidad de macollos
Mathew et al. (1995) demostraron que la forma de relación entre tamaño y densidad de macollo difiere con el IAF y con la morfología del macollo, ocurriendo una compensación entre tamaño y densidad de macollo cuando se alcanza el potencial ambiental. Este concepto no es fácilmente aplicable cuando se trabaja con pastoreos con carga intermitente, en que las variaciones en el IAF son máximas (Lemaire y Chapman, 1996).
Se encontró relación entre ambas variables en dos fechas. El 22 de octubre la función ajustada significativa (P<0,011) fue cuadrática con un r2=0,76 (Figura 8). La ecuación obtenida: Peso de macollo (mg)=-963,9+0,58 x -0,000079 x2, siendo x=número de macollos/m2, indica que en el rango menor de densidad de macollos (2700 a 3400 macollos/m2) la relación con el peso de los mismos es positiva. Si las densidades son bajas o no hay competencia por luz, puede darse un aumento en la contribución de materia seca por un aumento del tamaño de los macollos y/o en la densidad de los mismos.
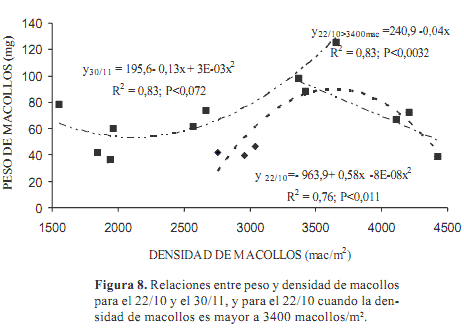
Pero desde 3.400 macollos/m2 cada incremento en el número de macollos significa una disminución en el peso de los mismos (partiendo de 98 mg/macollo) dándose el mecanismo de compensación mencionado. A partir de este punto, la ecuación lineal de disminución del peso de los macollos al aumentar la densidad es: Peso de macollo (mg)=240,9-0,04x, con una P<0,0032, y un r2=0,83.
Existió también una relación significativa (P<0,011) entre ambas variables al final del período experimental (30 de noviembre). La función cuadrática, Peso de macollo (mg)=195,6–0,13x + 0,000032x2, con un r2=0,83, fue inversa, manifestando que los cambios ocurridos en la estructura de la pastura al final de la primavera no estuvieron dirigidos por una limitación de luz. Seguramente afectó la limitada disponibilidad de energía para el rebrote.
Podemos expresar que la relación tamaño/densidad evolucionó en el tiempo y no alcanzó su equilibrio final, mostrando un modelo multifásico como el reportado por Matthew et al. (1995). Al final del período experimental se observó una reducción en la densidad y peso de los macollos, indicando que las condiciones ambientales y de manejo no fueron óptimas para la producción y sobrevivencia de los macollos.
Relaciones con el forraje remanente
El número y peso de los macollos residuales al pastoreo y su contenido de carbohidratos de reserva, determinan el rebrote (Fulkerson, 1994). Lee et al. (2008) encontraron una relación lineal entre las defoliaciones a cinco alturas diferentes (de 2 a 10 cm) y el contenido de carbohidratos de reserva. La altura de la defoliación también actúa a través de cambios en los requerimientos de las plantas (Fulkerson y Slack, 1995). A menor área foliar residual, el rebrote se realiza a partir de tejidos menos diferenciados y por lo tanto a menores tasas de crecimiento iniciales (Grant et al., 1981), determinando macollos más livianos si los intervalos entre pastoreos son menores al óptimo (Fulkerson y Slack, 1995).
En este caso la relación de la altura del forraje remanente pos pastoreo con las OF fue significativa (P<0,0001) y lineal. La ecuación para el período fue: Altura del forraje remanente (cm)=1,2+1,2 OF, con un r2=0,74.
El peso por macollo del forraje remanente también presentó una relación lineal significativa (P<0,0001) con un r2=0,83, incrementando 4,8 mg por cada 1 % de incremento en la OF a partir de 2,0 mg.
Únicamente para el 22 de octubre se encontró una relación significativa entre la densidad de macollos pre pastoreo con el peso de los macollos remanentes del pastoreo anterior (P<0,0037), con un r2=0,78, siendo Densidad de macollos pre pastoreo=2412,9+30,4 x, para x=peso de macollo del forraje remanente del pastoreo del 20 de septiembre.
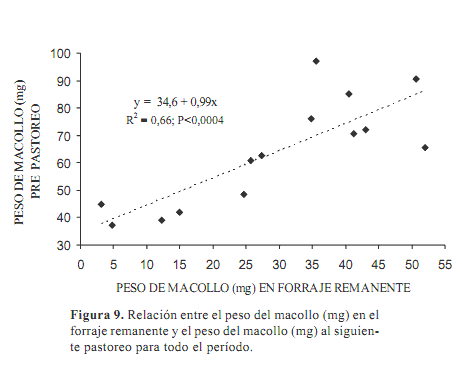
La relación entre el peso promedio de los macollos pre pastoreo con el peso de los macollos remanentes del pastoreo anterior (Figura 9) fue significativa (P<0,0004), con un r2=0,66 para todo el período estudiado. La ecuación fue: Peso de macollo pre-pastoreo (mg)=34,6+1,0 x, siendo x=peso de macollo del forraje remanente del pastoreo anterior.
Conclusiones
Las diferentes intensidades de pastoreo logradas a través de las OF provocaron variaciones en las variables estructurales estudiadas: densidad de macollos, densidad de plantas, número de macollos/planta y peso de macollos. Las relaciones halladas difirieron con la fecha y/o número de pastoreos realizados, ya que las condiciones ambientales fueron cambiando así como el estado de desarrollo de los macollos (pasaje al estado reproductivo).
También existieron diferencias en el momento en que comenzaron a detectarse las relaciones, así como diferentes comportamientos de las variables según fueran macollos grandes o pequeños. Esto permitió visualizar cómo el trabajar con los valores promedio de la población no permite, muchas veces, comprender los procesos que suceden.
A mayor intensidad del pastoreo el tamaño de los macollos fue menor, pero esto no fue acompañado de un incremento en el número de macollos, sino al contrario. La limitante principal en el promedio del período no fue la luz sino el balance de fotosintatos. En los tratamientos más intensos con área foliar remanente menor o nula, disminuyó el número de macollos, seguramente debido a que la disponibilidad de energía para la refoliación fue limitada, lo que se deduce de las relaciones encontradas con la altura y peso/macollo del forraje remanente.
Las respuestas a las OF fueron más rápidas en el peso que en el número de los macollos, coincidiendo con las distintas vidas medias de los órganos (hojas vs tallo) y prioridades de la planta.
En nuestras condiciones no se llegan a alcanzar los IAF óptimos citados en la literatura ya que otros recursos son limitantes. En este trabajo el contenido de N en el suelo seguramente lo fue y determinó la magnitud de las respuestas. Por esto las densidades de los macollos logradas fueron sensiblemente menores a las de Reino Unido, determinando que la relación tamaño/densidad de macollos con la intensidad del pastoreo difiriera de otros resultados reportados.
Las relaciones halladas demuestran la complejidad de los procesos que determinan el rendimiento final de las pasturas (peso y número de macollos) y señalan la necesidad de continuar estudiándolos cuando el pastoreo es intermitente y las condiciones ambientales son variables y limitantes.
Agradecimientos
Se agradece a los Profesores Álvaro Simeone y Ramiro Zanoniani, responsables del proyecto del cual surgió este trabajo, y a los numerosos estudiantes de tesis que trabajaron en el mismo.
Bibliografía
Altamirano, A.; Da Silva, H.; Durán, A.; Echevarría, A.: Panario, D. y Puentes, R. 1976. Carta de reconocimiento de suelos del Uruguay. Instituto Geográfico Militar. Mdeo. Uruguay. 96 p.
Agnusdei, M. G. and Assuero, S.G. 2004. Leaf tissue flows under grazing and sward structure of different temperate forage grasses in the humid pampas of Argentina. En: «II Symposium in Grassld. Ecophysiology and Grazing Ecology». Curitiba. Paraná. Brasil.
Bahmani I.; Hazard L.; Varlet-Grancher C.; Betin M.; Lemaire G.; Matthew C. and Thom E.R. 2000. Differences in tillering of long and short leaved perennial ryegrass genotypes under full light and shade treatments. Crop Sci. 40:1095-1102.
Bircham, J. S. and Hodgson, J. 1983. The influence of sward condition on rates of herbage growth and senescence in mixed swards under continuous stoking management. Grass Forage Sci. 38:323-331.
Briske, D.D. 1986. Plant response to defoliation: morphological considerations and allocation priorities. En: Rangelands: A resource under siege. Eds. Joss, P.J.; Lynch, P.W., Williams, O.B. Cambridge. Univ. Press. pp. 425-427.
Brock, J.L. and Fletcher, R.H. 1993. Morphology of perennial ryegrass (Lolium perenne) plants in pastures under intensive sheep grazing. J. agric. Sci., Camb. 120:301-310.
Calsal, J.J.; Sanchez, R.A. and Gibson D. 1990. The significance of changes in the red/far red ratio, associated with either neighbour plants or twilight, for tillering in Lolium multiflorum Lam. New Phytol.116:565-572.
Donaghy, D. J. and Fulkerson, W. J. 1997. The importance of water-soluble carbohydrate reserves on regrowth and root growth of Lolium perenne (L.). Grass Forage Sci. 52:401-407.
Donaghy, D.J. and Fulkerson, W. J. 1998. Priority for allocation of water-soluble carbohydrate reserves during regrowth of Lolium perenne. Grass Forage Sci. 53:211-218.
Duru, M. and Ducrocq, H. 2000. Growth and senescence of the successive grass leaves on a tiller. Ontogenic development and effect of temperature. Ann. Bot. 85:635-643.
Fulkerson, W.J. 1994. Effect of redefoliation on the regrowth and water soluble carbohydrate content of Lolium perenne. Aust. J. Agric. Res. 45:1809-1815.
Fulkerson, W.J. and Slack, K. 1994. Leaf number as a criterion for determining defoliation time for Lolium perenne: 1. Effect of water-soluble carbohydrates and senescence. Grass Forage Sci. 49:373-377.
Fulkerson, W.J. and Slack, K. 1995. Leaf number as a criterion for determining defoliation time for Lolium perenne: 2. Effect of defoliation frecuency and height. Grass Forage Sci. 50:16-20.
Fulkerson, W.J. and Donaghy, D.J. 2001. Plant-soluble carbohydrate reserves and senescence key criteria for developing an effective grazing management system for ryegrass-based pastures: a review. Aust. J. Exp. Agric. 41:261-275.
García, G.; García Pintos, L. y López; M. 2008. Efecto de la suplementación energética sobre la performance de novillos manejados sobre una mezcla de raigrás perenne bajo cuatro presiones de pastoreo. Tesis Ing. Agr. Montevideo, Uruguay. Facultad de Agronomía. 189 p.
Gastal, F.; Lemaire, G. and Lestienne, F. 2004. Defoliation, shoot plasticity, sward structure and herbage utilisation. En: II Symposium in Grassld. Ecophysiology and Grazing Ecology. Curitiba. Paraná. Brasil.
Gautier, H.; Varlet-Grancher, C. and Hazard, L. 1999. Tillering responses to the light environment and to defoliation in populations of perennial ryegrass (Lolium perenne L.) selected for contrasting leaf length. Ann. Bot. 83:423-429.
Grant, S. A.; Barthram, G.T. and Torvell, L. 1981. Components of regrowth in grazed and cut Lolium perenne swards. Grass Forage Sci. 36:155-168.
Grant, S. A.; Barthram, G.T.; Torvell, L.; King, J. and Smith, H.K. 1983. Sward management, lamina turnover and tiller population density in continuously stocked Lolium perenne-dominated swards. Grass Forage Sci. 38:333-344.
Hernández Garay, A.; Hodgson, J. and Matthew, C. 1997. Effect of spring grazing management on perennial ryegrass and ryegrass-white clover pastures. 2. Tiller and growing point densities and population dynamics. N. Z. J. Agric. Res. 40:37-50.
Hernández Garay, A.; Matthew, C. and Hodgson, J. 2000. The influence of defoliation height on dry matter partitioning and CO2 exchange of perennial ryegrass miniature swards. Grass Forage Sci. 55:372-376.
Hepp, C.; Milne, J.A.; Illius, A.W. and Robertson, E. 1996. The effect of summer management of perennial ryegrass-dominant swards on plant and animal responses in the autumn when grazed by sheep. 1. Tissue turnover and sward structure. Grass Forage Sci. 51:250-259.
Hodgson, J. 1990. Grazing management: Science into practice. Eds. Whitttemore, Simpson. New York, USA. Longman Scientific Technical. 203 p.
Hume, D.E. and Brock, J.L. 1997. Morphology of tall fescue (Festuca arundinacea) and perennial ryegrass (Lolium perenne) plants in pastures under sheep and cattle grazing. J. agric. Sci., Camb. 129:19-31.
Jewiss, O.R. 1972. Tillering in grasses: Its significance and control. J. Br. Grassld. Soc. 27:65-82
Lee, J.M.; Donaghy, D.J. and Roche, J.R. 2008. Effect of defoliation severity on regrowth and nutritive value of perennial ryegrass dominant swards. Agron. J. 100:308-314.
Lemaire, G. and Chapman, D. 1996. Tissue flows in grazed plant commmunities. En: The Ecology and Management of Grazing Systems. Eds. Hodgson, J.; Illius, A.W. Wallingford, UK. CAB International, pp.3-35
Lemaire, G. 2001. Ecophysiology of Grassland: Dynamic Aspects of Forage Plant Populations in Grazed Swards. En: Proc. XIX Int. Grassld. Cong., Sao Paulo, Brasil.CD.
Matthew, C.; Lemaire, G.; Sackville Halmiton, N.R. and Hernández-Garay, A. 1995. A modified self-thinning equation to describe size/density relationships for defoliated swards. Ann. Bot. 76:579-587.
Matthew, C.; Assuero, S.G.; Black, C.K. and Sackville Halmiton, N.R. 2000. Tiller dynamics of grazed swards En: Grassland Ecophysiology and Grazing Ecology Eds. Lemaire, G; Hodgson, J.; De Moraes, A.; Nabinger, C.; Carvalho, P.C. de F. Oxford, CABI. Publishing, pp.127-150.
Ong, C. K. 1978. The physiology of tiller death in grasses. 1. The influence of tiller age, size and position. J. Br. Grassld. Soc. 33:197-203.
Parsons, A.J. and Chapman, D.F. 2000. The principles of Pasture Growth and Utilization. En: Grass: Its production and utilization, Ed. Hopkins, A., Blackwell Science, Oxford. pp.31-89.
Peacock, J.M. 1975. Temperature and leaf growth in Lolium perenne.III. Factors affecting seasonal differences. J. Appl. Ecol. 12:685-697.
Santibáñez, F. 1994. Zonas Agroclimáticas de América del Sur. Universidad de Chile, Escuela de Agronomía, Laboratorio de Agrometeorología (mapa).
SAS. Institute Inc., SAS/STAT. User’s guide, Versión 8.1, Carey, North.Caroline, U.S.A.
Skinner, R.H. and Nelson, C. J. 1994. Role of leaf appearance rate and the coleoptile tiller in regulating tiller production. Crop Sci. 34:71-75.
Thornthwaite, C.W. and Mather, J.R. 1955. The water balance. Centerton, N. J. Laboratory of Climatology.

