










Serviços Personalizados
Journal

Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Links relacionados
Compartilhar

 Permalink
PermalinkArchivos de Medicina Interna
versão impressa ISSN 0250-3816versão On-line ISSN 1688-423X
Arch. Med Int vol.37 no. 1 Montevideo mar. 2015
Revisión
Importancia de la evaluación del estrés oxidativo en el semen humano
Importance of the assessment of oxidative stress in human semen
Bioquím. Cecilia Vicenta Paparella
Bioquímica del Laboratorio Estudios Especiales en Semen. Profesora del Área Química Analítica Clínica. Facultad de Ciencias Bioquímicas y Farmacéuticas. Universidad Nacional Rosario. Argentina.
Bioquím. Adriana Beatriz Pavesi
Bioquímica del Laboratorio Estudios Especiales en Semen. Profesora del Área Química Analítica Clínica. Facultad de Ciencias Bioquímicas y Farmacéuticas. Universidad Nacional Rosario. Argentina.
Dr. Rodolfo Néstor Feldman
Médico Andrólogo del Servicio de Reproducción de la Facultad de Ciencias Médicas. Universidad Nacional Rosario. Argentina.
Bioquím. Beatriz Reina Bouvet
Bioquímica del Laboratorio Estudios Especiales en Semen. Profesora del Área Química Analítica Clínica. Facultad de Ciencias Bioquímicas y Farmacéuticas. Universidad Nacional Rosario. Argentina.
Recibido: 04/09/14 - Aceptado: 10/02/15
Correspondencia: Cecilia Vicenta Paparella - Ovidio Lagos 371 (2000) Rosario.
Tel.: 54-0341-155859224. e-mail: ceciliapaparella@yahoo.com.ar
RESUMEN: Arch Med Interna 37(1): 07-14
Utilizando bases de datos Medline, Science Direct, Endocrine Society y Sociedad Argentina de Andrología se realizó una revisión bibliográfica para estudiar e interpretar el efecto del estrés oxidativo (EO) en el proceso reproductivo. Los seres vivos que utilizan oxígeno para obtener energía son liberadores de especies reactivas de oxígeno (ERO). Existen situaciones andrológicas, factores medioambientales y compuestos químicos que incrementan las citoquinas proinflamatorias, generan EO alterando la regulación del proceso espermatogénico. Los espermatozoides son susceptibles al daño oxidativo, debido al elevado contenido de ácidos grasos poliinsaturados (AGPI) en su membrana y el escaso citoplasma que limita el contenido de enzimas antioxidantes. El EO induce daño peroxidativo en la membrana espermática y fragmentación del ADN en los genomas nucleares y mitocondriales. El test estrés espermático modificado (MOST) estima la resistencia espermática a la lipoperoxidación. El incremento de ERO en hombres con trastornos reproductivos y su correlación con alteraciones seminales revelan la importancia de la evaluación del EO en el estudio integral del hombre que consulta por infertilidad.
Palabras clave: Estrés oxidativo - Proceso reproductivo - Especies reactivas de oxígeno - Espermatozoide - Infertilidad masculina.
Abstract: Arch Med Interna 37(1): 07-14
Data from Medline, Science Direct, Endocrine Society and organisms of information of Argentina Society of Andrology were used to study and interpret the effect of oxidative stress (OS) in the reproductive process. The living things use the oxygen for energy they release reactive oxygen species (ROS). There andrology situations enviromental factors, chemical agents that increase of proinflammatory cytokines, OS generated by altering the regulation of spermatogenesis process. The spermatozoids due to high concentration of polyunsaturated fatty acids (PFA) in its membrane are sensitive to the oxidative damage induced for ROS and it also contain low levels of antioxidant enzymes for their limited cytoplasm. OS induced peroxidative damage in sperm membrane and DNA fragmentation in the nuclear and mitochondrial genomes. The modified sperm stress test (MOST) estimates sperm resistance to lipoperoxidation. The ROS increased in men with reproductive disorders and its correlation with seminal alterations reveal the importance of the evaluation of OS in the study of man consulting for infertility.
Keywords: Oxidative stress - Reproductive process - Reactive oxygen species - Spermatozoid - Men infertility.
INTRODUCCIÓN
La infertilidad se define como la incapacidad de una pareja para concebir luego de un año de relaciones sexuales sin medidas anticonceptivas. Alrededor del 20% de las parejas en edad reproductiva tienen problemas de infertilidad. El factor masculino está involucrado en alrededor del 50% de las parejas infértiles, 30% sólo por factor masculino y 20% son causas compartidas(1). En cuanto al origen genético de la infertilidad masculina actualmente se considera menor al 5%(2). Por otra parte un 10 a 15% de los varones infértiles presentan parámetros seminales dentro de los rangos de referencia. En estos casos las causas de infertilidad masculina podrían ser defectos estructurales del núcleo o de la membrana espermática, factores ambientales, ocupacionales y además genéticos(3).
La evaluación inicial del hombre en una pareja infértil incluye un examen clínico integral, con particular énfasis en la historia clínica del sistema endócrino-reproductor. El examen del semen realizado según los lineamientos propuestos por la Organización Mundial de la Salud (OMS), es la piedra angular para la evaluación de la capacidad reproductiva del varón y constituye una herramienta esencial al comenzar a estudiar la pareja que consulta por infertilidad(4).
La función espermática se investiga mediante un grupo de determinaciones, donde cada prueba está dirigida a una organela diferente. Se emplean test con alta especificidad, precisos y objetivos, que contribuyen al conocimiento de una disfunción reproductiva en particular. Además el laboratorio andrológico cuenta con una batería de ensayos que estudian in vitro los diferentes pasos de la función espermática desde la migración hasta la unión del espermatozoide a la zona pelúcida, la fusión espermatozoide-ovocito y la decondensación de la cromatina(5).
En la capacitación espermática, las ERO también juegan un papel decisivo, se ha comprobado que bajo condiciones fisiológicas y de forma controlada, la producción de ERO es de vital importancia para la adquisición de la capacidad fecundante del espermatozoide(6). Por otra parte, se conoce que las ERO pueden incidir negativamente sobre la fertilidad masculina a través de dos mecanismos fundamentales. El primero de ellos es el daño que provocan a la membrana espermática, lo cual da lugar a una reducción de la movilidad y habilidad del espermatozoide de fusionarse con el óvulo. En tanto, el daño que las ERO ejercen directamente sobre el ADN espermático, es otro de los mecanismos a través del cual afectan la genómica paternal del embrión. La integridad del ADN constituye un indicador de la espermatogénesis y de la fertilidad masculina(7).
En casos de infertilidad masculina idiopática es frecuente encontrar en el semen elevadas concentraciones de ERO, en la inactivación de las mismas participan los agentes antioxidantes presentes en el fluido seminal. Cuando este equilibrio se pierde genera EO que conlleva a desregulaciones metabólicas y funcionales que provocan muerte celular e infertilidad(8).
ESPECIES REACTIVAS DE OXÍGENO
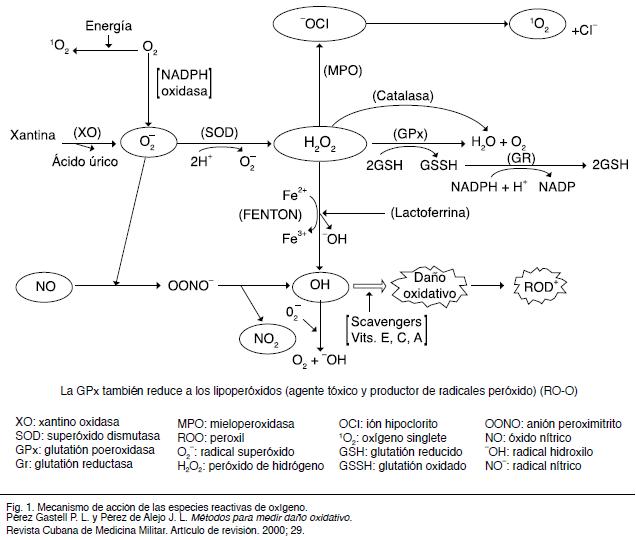
Las ERO incluyen iones de oxígeno, radicales libres y peróxidos, tanto inorgánicos como orgánicos. Se forman de manera natural como subproducto del metabolismo normal del oxígeno y desempeñan un papel importante en la señalización celular. Todos los seres vivos que utilizan el oxígeno para obtener energía liberan ERO. Las células poseen mecanismos de defensa denominados genéricamente como antioxidantes (figura 1).
El espermatozoide dispone de mecanismos enzimáticos como:
• superóxido dismutasa (SOD), capta el radical anión superóxido (O2-) mediante una dismutación para convertirlo en peróxido de hidrógeno (H2O2);
• glutatión peroxidasa/glutatión reductasa, neutralizan al H2O2;
• citocromo oxidasa, evita la reducción univalente del oxígeno.
El plasma seminal es una potente fuente de antioxidantes compuesta por:
• enzimas antioxidantes de alto peso molecular: SOD, glutatión peroxidasa/glutatión reductasa, catalasa;
• antioxidantes de bajo peso molecular: tocoferol o vitamina E, neutralizan al radical oxhidrilo (OH) y se encuentran en las membrana;
• vitamina C, es un reductor que reacciona rápidamente con el oxígeno y con el OH, también capta el oxígeno singlete (O) y el ión hipoclorito;
• glutatión, puede captar el H2O2, O y OH;
• albúmina, piruvato, β-carotenos, taurina, ácido úrico.
Los niveles patológicos de ERO detectados en el semen de hombres infértiles generalmente se deben a un incremento en la producción y no a una reducción en la capacidad antioxidante del plasma seminal. Los espermatozoides debido al alto contenido de AGPI son susceptibles al daño inducido por las ERO. Estudios recientes demostraron que las mitocondrias son generadoras de EO, previo al desarrollo de la cascada apoptótica generan elevados niveles de anión O2- cuyo efecto oxidativo se incrementa por la incompetencia del espermatozoide para contrarrestar el daño oxidativo debido a su escaso citoplasma y los bajos niveles de enzimas antioxidantes(9). Uno de los efectos de las ERO es la alteración de la membrana celular mediante peroxidación lipídica (PL). Este proceso fisiopatológico tiene como resultado una cascada de profundos cambios degradativos que afectan la organización y función de la membrana del espermatozoide humano, producen rupturas del ADN y degradación proteica. En la etiología de la infertilidad masculina, un factor significativo es el que involucra la pérdida de la función espermática como consecuencia del EO, el cual puede provenir de la deficiencia del sistema de defensa antioxidante o del incremento de la formación de ERO(10).
Durante el desarrollo testicular, se producen cambios de las enzimas antioxidantes presentes en el testículo y el EO que se produce durante el proceso espermatogénico induce daño peroxidativo en la membrana plasmática espermática y fragmentación del ADN en los genomas nucleares y mitocondriales. Por otra parte el espermatozoide humano, requiere de niveles bajos de ERO para su normal funcionamiento y sufre procesos controlados de óxido-reducción durante la hiperactivación espermática, capacitación y reacción acrosomal(11). En la capacitación espermática también intervienen otros mecanismos que están regulados por ERO como es el influjo de calcio a través de la membrana, el aumento del adenosin monofosfato cíclico (AMPc) y la fosforilación de proteínas(12).
Macrófagos y granulocitos activados producen H2O2, O2-, nitratos y nitritos que son altamente tóxicos para el espermatozoide porque inducen el daño peroxidativo. Varios autores observaron que los leucocitos presentes en el semen pueden causar EO, incluso a concentraciones inferiores del límite de corte establecido por la OMS(4,10).
Mecanismo de daño espermático mediado por ERO
Las ERO pueden dañar proteínas, ácidos nucleicos y causar pérdida de la movilidad espermática afectando la fertilidad. Debido al alto contenido de AGPI, los espermatozoides humanos son especialmente sensibles al daño por ERO. Los lipoperóxidos y sus productos de degradación como el malondialdehído (MDA) y los 4-hidroxialquenales (4-HA) son altamente tóxicos para las células sexuales masculinas y provocan daños irreversibles en la movilidad. La PL está correlacionada con la disminución de la movilidad espermática y con los defectos morfológicos de las gametas masculinas(13). Este fenómeno consiste en el daño oxidativo sobre los AGPI, los cuales contienen en su estructura dos dobles enlaces C-C especialmente susceptibles a la sustracción de un electrón por parte de las ERO. Además conduce a un aumento considerable de la permeabilidad de las membranas celulares, y origina alteraciones irreversibles de las propiedades funcionales de la célula que pueden conducir a su lisis total(14-16).
Durante la fase de iniciación de la PL, un radical de elevada reactividad como el OH- puede extraer un átomo de hidrógeno de un ácido graso insaturado para producir el radical lipídico, el cual se reorganiza y forma el dieno conjugado que en su reacción con el O2 da lugar al radical peroxilo con suficiente reactividad como para atacar otro lípido y conducir a la propagación de la reacción oxidativa(17). La conjugación rápida de dienos con O2 favorece la formación de numerosos derivados tóxicos como son los hidroperóxidos (ROOH), los 4-HA y el MDA(18).
Las ERO pueden causar hipercondensación del núcleo espermático como resultado de una excesiva oxidación de los grupos sulfhidrilos proteicos. Aproximadamente el 10% de los espermatozoides provenientes de hombres fértiles poseen niveles medibles de daño al ADN, mientras que en aquellos que provienen de hombres infértiles, este porcentaje oscila entre 20 y 25%(16). Las ERO son capaces de dañar el ADN a través de la oxidación de las bases de purina y pirimidina, así como del esqueleto carbonado. Normalmente el ADN se encuentra empaquetado y protegido por las protaminas, en la infertilidad masculina se conoce que existe un déficit de estas proteínas(19). Otro de los daños que provocan es la activación de la apoptosis a través de la degradación enzimática del ADN por acción de las caspasas(20). Aunque la mayor parte del ADN espermático es nuclear, existe una pequeña fracción que es de origen mitocondrial. Este ADN codifica para 37 genes, de los cuales 13 lo hacen para polipéptidos que forman parte de subunidades de los complejos enzimáticos involucrados en el proceso de fosforilación oxidativa. Ésta puede ser una de las causas por las cuales el espectro de mutación del ADN mitocondrial en el espermatozoide puede ser de 10 a 100 veces mayor que el ADN nuclear(21).
Evaluación de ERO
El daño oxidativo se puede medir de forma directa o indirecta.
Métodos directos: se ha tratado de medir la concentración de agentes oxidantes en el organismo, pero es dificultoso debido a que tienen una vida media muy corta. Las técnicas de quimioluminiscencia aplicadas en la detección de ERO han incorporado la peroxidasa del rábano para aumentar la eficiencia de la reacción. Estos ensayos utilizan un luminómetro a 37ºC, se exponen los espermatozoides a una solución de luminol en dimetil sulfóxido y se monitorea la señal quimioluminiscente. La medición de ERO es sencilla pero requiere de luminómetros muy sensibles capaces de medir señales quimioluminiscentes de metabolitos oxígeno reactivos(22).
Métodos indirectos: Evalúan las ERO midiendo los productos terminales de su acción oxidante sobre las proteínas, los lípidos y el ADN. Para cuantificar los compuestos que reaccionan con el ácido tiobarbitúrico (ATB) se aplica un método espectrofotométrico que mide la generación de MDA como mayor producto del desdoblamiento de los hidroperóxidos estimulado con hierro y ascorbato en presencia de ATB. El hierro induce una rápida ruptura de los peróxidos preexistentes en el plasmalema, por lo que la medida de la peroxidación es un indicador del grado de EO experimentado por una población de espermatozoides. Este método tiene poca sensibilidad y la conversión a MDA no es total(22).
El MOST estima la resistencia de los espermatozoides a la lipoperoxidación. Este ensayo funcional mide los cambios en la movilidad espermática luego de incubar los espermatozoides móviles, obtenidos post swim-up, a temperatura de 40°C. El MOST permite además predecir fallas de la fertilización de origen espermático sobre ovocitos humanos y se aplica en prácticas de reproducción asistida. El valor de corte del ensayo MOST se estableció en 0,40 porque presenta índices máximos de sensibilidad, especificidad, valores predictivos positivos y negativos de la prueba. Los niveles bajos de MOST (< 0,40) reflejan un aumento de la PL que altera la capacidad fertilizante del varón(23).
Efectos de ERO en la membrana plasmática
La membrana espermática es asimétrica en su estructura y en sus funciones. Está formada por una asociación de fosfolípidos y esteroles en equilibrio dinámico con proteínas de membrana. El colesterol y los fosfolípidos son importantes en el mantenimiento de la integridad estructural de los sistemas de membrana. Por otra parte el elevado contenido de fosfolípidos unidos a ácidos grasos insaturados incrementa la vulnerabilidad espermática a los cambios peroxidativos(24).
El colesterol es muy abundante en espermatozoides humanos, se encuentra en la bicapa de fosfolípidos, reduce la permeabilidad de la membrana y limita la inserción de proteínas. La disminución del colesterol de la membrana plasmática constituye un paso importante en la capacitación espermática. La integridad y funcionalidad de la membrana es esencial para el proceso de capacitación, reacción acrosomal, unión a la zona pelúcida y al oolema. Se evalúa por la capacidad de permitir el transporte de determinados fluidos y ciertas moléculas a través de ella. El test hipoosmótico (THO) mide la capacidad de los espermatozoides para incorporar agua especialmente a nivel del segmento intermedio y de la cola cuando son incubados a 37ºC en un medio hipoosmótico (150 mOsm/l). Los resultados del THO se correlacionan con la morfología, la movilidad y la vitalidad espermáticas(4). Se estudió en muestras de semen de pacientes infértiles, la relación del EO con la funcionalidad de la membrana espermática observando asociación estadísticamente significativa entre las muestras con MOST anormal y resultados alterados del THO(24,25).
Efecto de ERO en el ADN espermático
La integridad de la estructura de la cromatina espermática depende de factores endógenos como la maduración espermática y exógenos como el daño peroxidativo del ADN, infecciones, factores inmunológicos o diversos agentes químicos(26).
Durante la espermiogénesis las histonas, proteínas clásicas de empaquetamiento del ADN, son reemplazadas por proteínas de transición y luego por proteínas más básicas ricas en residuos de arginina y cisteína denominadas protaminas. Durante la maduración epididimaria de la gameta masculina las protaminas participan en la formación de puentes disulfuro intra e intermoleculares proporcionando mayor estabilidad, protección y compactación de la cromatina, constituyendo una estructura química, mecánicamente resistente y transcripcionalmente inactiva(4).
Los cambios en la cromatina nuclear protegen al genoma paterno durante el transporte del espermatozoide por el tracto reproductor masculino y femenino. Las fallas en el proceso de protaminación están relacionadas con la apoptosis espermática, la fragmentación de las cadenas del ADN y las anomalías cromosómicas(27).
Los defectos del núcleo espermático son de gran significación en infertilidad masculina. Se han descripto vacuolas intranucleares que contienen inclusiones pleomorfas, como grupos de membranas, vesículas, gotas de lípidos, organelas citoplasmáticas que asociadas con anormalidades en la condensación de la cromatina producen menor tasa de embarazos. La identificación ultraestructural y la cuantificación de estas alteraciones permiten adoptar las medidas terapéuticas tendientes a corregir el factor andrológico involucrado o arbitrar métodos de selección con microscopía de alta resolución en los procesos de microinyección(28).
El test azul de anilina (TAA) evalúa el estado de condensación de la cromatina y se fundamenta en la tinción selectiva de las proteínas ricas en lisina. La presencia de espermatozoides cuyos núcleos están teñidos con azul de anilina indica deficiencia en la protaminación nuclear(4). El 10-15% de los ovocitos no fertilizados mediante procedimiento de inyección intracitoplasmática (ICSI) presentan espermatozoides con cromatina inmadura y el empaquetamiento incorrecto de la cromatina podría ser la causa del fracaso en la fertilización(28).
Fragmentación del ADN espermático
La fragmentación del ADN consiste en interrupciones en las cadenas simples o dobles. La transferencia de la molécula de ADN integra e intacta desde el espermatozoide al óvulo es esencial para conseguir una fecundación con ciertas perspectivas de éxito. Una ruptura podría llevar a alteraciones en la fertilización, en el desarrollo embrionario y fetal e incluso puede afectar la salud de la descendencia. Aproximadamente el 25% de hombres infértiles tienen elevado porcentaje de fragmentación del ADN. Este daño es multifactorial y puede producirse en cualquier etapa del proceso espermatogénico(29).
Altos niveles de ERO alteran la movilidad y la morfología espermática, provocan fragmentación del ADN que puede acelerar el proceso de apoptosis de las células germinales, disminuyendo la concentración espermática con el consecuente deterioro de la calidad seminal(8).
La fragmentación del ADN espermático inducida por el radical OH resulta en la formación de 8-OH-guanina y 8-OH-2´ deoxiguanosina en un primer estadio seguido de fragmentación del ADN de cadena doble. Mientras que el daño del ADN en el primer estadio podría ser reparado por el ovocito, la fecundación del ovocito por un espermatozoide con fragmentación del ADN de cadena doble es prácticamente irreversible e incompatible con el desarrollo normal del embarazo. La magnitud del daño inducido por radicales libres durante el tránsito espermático por el epidídimo depende de los niveles de radicales libres producidos por espermatozoides inmaduros, la presencia de células epiteliales o de leucocitos activados en el epidídimo y de los niveles de enzimas antioxidantes presentes en la luz del epidídimo(30).
Los espermatozoides más vulnerables son los que poseen menor grado de entrecruzamiento de puentes disulfuro en la cromatina. La fragmentación del ADN en los espermatozoides eyaculados es más elevada que en los espermatozoides testiculares o epididimarios. El epidídimo posee un mecanismo selectivo que elimina los espermatozoides genómicamente defectuosos. Por otra parte el tratamiento farmacológico con antioxidantes reduce significativamente los niveles de fragmentación del ADN espermático(31).
La caracterización del daño al ADN a nivel celular es importante, si bien el ovocito está capacitado para reparar el ADN del espermatozoide que lo fecunda, este proceso es más difícil cuando se trata de alteraciones del ADN cadena doble y además depende de la calidad genómica y citoplasmática del ovocito. En general el factor genómico de infertilidad incluye el daño del ADN espermático, alteraciones de la meiosis y aneuploidías en espermatozoides, de los cuales sólo el daño del ADN es reparado por el ovocito(32).
Métodos para evaluar la fragmentación del ADN
Por varios años los investigadores han buscado la mejor forma de cuantificar el daño presente en el ADN de espermatozoides humanos. Actualmente se dispone de varias técnicas que miden el daño al ADN espermático. Algunas evalúan la integridad del ADN y otros ensayos identifican defectos en el empaquetamiento de la cromatina.
Las que se aplican con mayor frecuencia en los laboratorios andrológicos y de reproducción son:
• Test de heterogeneidad de la cromatina (TNA);
• Test de dispersión de la cromatina (SCD);
• Test Terminal dUTP Nick-End Labeling (TUNEL).
El TNA se fundamenta en la capacidad del fluorocromo naranja de acridina de intercalarse entre las dos cadenas de ADN como un monómero que al ser excitado emite color verde, mientras que si está unido al ADN cadena simple emite coloración anaranjada. La prueba se cuantifica mediante el cambio metacromático de color verde del ADN nativo al anaranjado cuando el ADN está desnaturalizado(3,4,28). En muestras de semen de pacientes con infertilidad idiopática se estudió el efecto del EO sobre el núcleo espermático, encontrándose mayor alteración en la integridad del ADN y cromatina inmadura en las muestras con MOST alterado(25).
La prueba SCD se basa en dos fenómenos: el primero es que las hebras de ADN que contienen rupturas son más fáciles de ser desnaturalizadas. De hecho, los extremos de las rupturas, que pueden ser de cadena sencilla o de cadena doble, son los orígenes de la desnaturalización cuando éstos se enfrentan a un tratamiento de carácter ácido. El segundo fenómeno es que los bucles compactados de fibra de cromatina nuclear, compuestos de ADN y proteínas, se desempaquetan cuando se extraen proteínas nucleares. Este proceso de relajación de los bucles de cromatina tras un tratamiento desproteinizante da lugar a halos periféricos de cromatina/ADN que emanan del residuo nuclear central(26,29).
El Test de TUNEL (Terminal desoxi-nucleotidil-Uridin-trifosfato, dUPT, deoxynucleotidil transferase Nick-End Labelling assay) permite visualizar la incorporación de nucleótidos marcados en los extremos de las rupturas existentes en el DNA, bien sean de cadena simple o doble. La reacción se cataliza, in situ, mediante la acción de una transferasa terminal. Esta enzima incorpora deoxiuridina modificada con biotina o digoxigenina, en el extremo 3’- OH de la cadena afectada. El nucleótido incorporado esta directamente marcado con un fluorocromo. La señal de marcado obtenida por cada espermatozoide, se incrementa de acuerdo con el número de rupturas que presente la cadena de DNA. Esta técnica ha tenido buena aceptación dado que es versátil, está comercializada en kit y los resultados pueden ser interpretados mediante microscopia de fluorescencia y citometría de flujo. Actualmente posee un valor de corte entre 15 y 30% según la metodología empleada(31,32).
Capacitación espermática y ERO
La capacitación espermática es un evento asincrónico. En humanos solamente el 10% de los espermatozoides están capacitados a un tiempo dado(12). Este proceso se asocia a:
• modificaciones en la composición y fluidez de la membrana plasmática;
• cambios en las concentraciones iónicas intracelulares;
• incremento en la fosforilación proteica;
• generación de niveles bajos y controlados de ERO.
La producción de anión O2- por los espermatozoides es un evento temprano en el proceso de capacitación. Las ERO regulan la actividad de enzimas implicadas en la síntesis de AMP cíclico y en la fosforilación de proteínas en residuos serina y treonina asociada con el eflujo de colesterol. Estos hallazgos manifiestan que numerosos cambios a nivel de la membrana plasmática junto con la activación de varias vías de señalización intracelular son necesarios para que el espermatozoide adquiera la capacidad de responder a los estímulos inductores de la exocitosis acrosomal(33).
ALTERACIONES SEMINALES ASOCIADAS A ERO
Oligozoospermia y criptorquidia
La oligozoospermia se caracteriza por una baja concentración de espermatozoides en el semen, se considerada moderada cuando es menor a 15 millones de espermatozoides/ml de semen y severa cuando es menor a 5 millones de espermatozoides/ml. La oligozoospermia idiopática es diagnosticada por exclusión de otras patologías o alteraciones, tales como varicocele, criptorquidia, obstrucción parcial de la vía seminal, presencia de anticuerpos antiespermáticos (AAE). Existen hallazgos clínicos y de laboratorio que se asocian a oligozoospermia, los cuales incluyen: edad, patología inflamatoria y autoinmune, disfunciones hormonales, ingesta de alcohol, factores genéticos y ambientales. Varios autores demostraron que los espermatozoides de pacientes con oligozoospermia idiopática o con antecedentes de criptorquidia, producen niveles elevados de ERO con incremento significativo en la extensión del daño en el ADN(34).
La criptorquidia es una patología congénita que afecta el 4 a 5% de niños recién nacidos. La exposición del testículo a la temperatura corporal altera el proceso espermatogénico, y además representa un factor de riesgo de cáncer testicular(35).
Varicocele
El varicocele es una dilatación anómala de las venas del plexo pampiniforme, las cuales desempeñan una función importante en el mantenimiento de la temperatura testicular mediante un mecanismo de intercambio contracorriente con la arteria espermática, manteniendo la temperatura gonadal aproximadamente en 35ºC. El éstasis venoso y la acumulación de desechos tóxicos que produce esta lesión vascular alteran la función testicular. Varios investigadores reportaron que el varicocele está asociado con elevados niveles de ERO producidos por el espermatozoide y disminución de la capacidad antioxidante del plasma seminal(36).
Otros autores con la finalidad de verificar si la determinación de ERO en semen tiene un valor predictivo sobre la probabilidad de desarrollar atrofia testicular y alteraciones espermáticas, estudiaron en pacientes con varicocele la asociación entre altos niveles de ERO, el grado de varicocele y el volumen testicular. Observaron asociación directa entre los niveles de ERO y el grado de varicocele pero no encontraron asociación con el grado de atrofia testicular(37).
Por otra parte estudios de la calidad y la funcionalidad espermática en hombres con varicocele, mostraron menor porcentaje de espermatozoides con movilidad progresiva, mayor porcentaje de gametas masculinas muertas, aumento de alteraciones en la morfología espermática y disminución del porcentaje de espermatozoides con membrana funcional. El varicocele altera parámetros seminales relacionados con la funcionalidad espermática deteriorando la capacidad reproductiva masculina(38).
FACTORES MEDIOAMBIENTALES ASOCIADOS A ERO
La contaminación medioambiental es un problema mundial. Anualmente se producen e incorporan al mercado más de 2.000 compuestos químicos de síntesis, que al ser liberados al medioambiente pueden interaccionar con los seres vivos ocasionando daños a la salud humana general y en especial a la salud reproductiva(39).
La exposición a diversos contaminantes medioambientales se asocia con la ocupación laboral, siendo las principales vías de ingreso al organismo la piel y la vía respiratoria. Muchas de estas sustancias químicas naturales o sintetizadas por el hombre, se comportan como hormonas alterando la homeostasis normal del sistema endocrino del individuo mediante un desequilibrio en el balance de estrógenos, andrógenos y hormonas tiroideas. Estos compuestos químicos se conocen como disruptores endocrinos y pueden unirse al receptor hormonal o antagonizar su efecto bloqueando al receptor correspondiente; en ambos casos alteran el sistema hormonal produciendo niveles elevados de ERO que generan daño celular(40).
El epitelio de los túbulos seminíferos del testículo donde se desarrolla la espermatogénesis es un medio rico en andrógenos, indispensables para la proliferación, diferenciación y maduración espermática. Toda alteración en el sistema hormonal masculino regulador de la función testicular, se manifiesta en modificaciones funcionales del espermatozoide. La carencia hormonal produce inmadurez en los espermatozoides y se manifiesta en el semen con elevada concentración de células germinales e incremento de las alteraciones morfológicas(41).
En una población de hombres con infertilidad idiopática se estudió el efecto de la exposición ocupacional a agroquímicos sobre parámetros seminales vinculados con la espermatogénesis. Se evaluaron trabajadores expuestos a agroquímicos y varones no expuestos como grupo control. El grupo de hombres expuestos presentó menor concentración espermática, mayor porcentaje de espermatozoides morfológicamente anormales y aumento marcado de células de la progenie espermática en el semen. La exposición laboral o accidental a agroquímicos altera el proceso espermatogénico deteriorando los parámetros seminales esenciales para el proceso de fertilización(42).
Por otra parte se evaluó la relación de la exposición ocupacional a solventes con la viabilidad espermática. Los solventes, ampliamente utilizados en la industria de la pintura y la madera, también se comportan como disruptores endocrinos. En este grupo de hombres ocupacionalmente expuestos se encontraron alteraciones en: la condensación de la cromatina, la integridad del ADN y la funcionalidad de la membrana espermática, estructuras espermáticas esenciales para la interacción con el ovocito(43).
Efecto del tabaco
El tabaco contiene sustancias tóxicas como la nicotina, que disminuyen los niveles séricos de gonadotrofinas. El humo del cigarrillo posee gran cantidad de hidrocarburos aromáticos policíclicos que actúan como inductores de enzimas, los que interfieren en la producción de esteroides sexuales y alteran el desarrollo armónico del proceso espermatogénico. Contiene además nitrosaminas y moléculas inorgánicas, muchas de ellas son sustancias reactivas del oxígeno y del nitrógeno. El aumento de ERO en hombres fumadores genera EO con incremento de citoquinas proinflamatorias que alteran la regulación de la espermatogénesis y la viabilidad espermática(44).
En hombres con infertilidad idiopática se estudió la relación del tabaquismo con parámetros seminales vinculados con el proceso espermatogénico: morfología espermática, concentración de espermatozoides y de células germinales inmaduras. Se analizaron 3 grupos de pacientes: fumadores de más de 20 cigarrillos/día, fumadores de menos de 20 cigarrillos/día y no fumadores como grupo control. El análisis estadístico de los resultados mostró diferencia significativa entre fumadores y no fumadores al comparar los promedios de las 3 variables analizadas. A diferencia de lo publicado por otros autores no se encontró diferencia significativa al relacionar la cantidad diaria de cigarrillos consumidos. El tabaquismo es un factor importante que debe ser evaluado al realizar el estudio integral del hombre infértil(45).
TRATAMIENTOS MÉDICOS PARA DISMINUIR EL IMPACTO DEL EO
Al plantear los tratamientos para reducir el EO es importante investigar las causas del desequilibrio oxidativo. Se demostró en individuos sanos que las dietas con altas concentraciones de antioxidantes están asociadas a elevada concentración de espermatozoides con buena movilidad espermática. Son beneficiosos los tratamiento con antioxidantes como vitaminas E y C, selenio, N-acetil-cisteína, glutatión y carnitina(46).
La L-carnitina (ácido 3-hidroxi-4-N-trimetil-aminobutírico) cumple un rol esencial en la producción de energía celular, además tiene efecto antioxidante sobre las membranas celulares y el ADN(47).
En pacientes con oligoastenoteratozoospermia de origen desconocido tratados con L-carnitina (2 g/día) más L-acetil-carnitina (500 mg/dos veces por día) durante 6 meses, se observaron incrementos en todos los parámetros espermáticos, siendo lo más significativo el aumento en la movilidad(48).
CONCLUSIONES
Todos los espermatozoides de un eyaculado no son iguales desde el punto de vista funcional, pero no se conoce cuáles son los atributos que diferencian un espermatozoide fértil de aquel que no lo es.
La incidencia mundial de la infertilidad masculina se ha incrementado notablemente en los últimos años debido a diversas causas, algunas de origen genético y otras asociadas a la exposición a agentes nocivos. El estudio de los mecanismos fisiopatológicos involucrados en la infertilidad masculina es de vital importancia para la aplicación de estrategias terapéuticas, ya que es conocido que la población mundial envejece al mismo tiempo que disminuye la natalidad(7).
Varios autores demostraron la correlación positiva entre la generación de anión superóxido mitocondrial y el contenido incrementado de AGPI presentes en la membrana de espermatozoides humanos senescentes y defectuosos(9). Estos espermatozoides alterados son particularmente vulnerables a la PL inducida por las mitocondrias espermáticas, la cual desencadena interacciones moleculares que provocan daño oxidativo conduciendo en forma inevitable a la pérdida de funcionalidad espermática y finalmente a la muerte celular(49). Si bien el ovocito es capaz de reparar el ADN dañado, la eficacia depende del grado de alteración nuclear y de la edad ovocitaria; no se conoce si el efecto que ejercen los factores medioambientales adversos, el tabaco y el estilo de vida sobre el ADN espermático, se correlaciona con la fracción de mutaciones presentes en la descendencia. Las técnicas de reproducción asistida facilitan la transferencia de ADN dañado. La susceptibilidad de los espermatozoides a la acción de ERO, la evidencia de niveles aumentados de EO en pacientes con trastornos reproductivos y su correlación con alteraciones en el análisis del semen y en la capacidad fecundante, sugieren la evaluación del EO en el estudio habitual de la pareja que consulta por infertilidad, con el fin de lograr un enfoque andrológico más racional y eficiente tendiente a instaurar medidas terapéuticas adecuadas(50-53).
BIBLIOGRAFÍA
1.Cruz L, Colmenares M, Berrueta-Carrillo L, Gómez-Pérez R, Montes H, Berrueta L et al. Evaluación de la calidad del espermatozoide humano: comparación entre la integridad del ADN espermático y variables del semen. Invest Clin 2010; 51 (1): 87-99.
2.Bhasin S. Approach to the infertile man. J Clin Endocrinol Metab 2007; 92: 1995-2004.
3.Reddy KV. Sperm membrane protein profiles of fertile and infertile men: identification and characterization of fertility-associated sperm antigen. Hum Reprod 2004; 19: 234-242.
4.WHO. 2010 Laboratory manual for the examination and processing of human semen. 5ºth ed.
5.Calamera J C, Calamera P. El factor espermático. CEGYR. Rev Med Reprod. 2004; 7 (1): 23-32.
6.De Lamirande E, Lamothe G. Reactive oxygen-induced reactive oxygen formation during human sperm capacitation. Free Radic Biol Med. 2009; 46: 502 – 510.
7.Tremellen K. Oxidative stress and male infertility-a clinical perspective. Hum Reprod Upd 2008; 14: 243-258.
8.Schteingart H F. Efecto de las especies reactivas de oxígeno en la infertilidad masculina. Aplicación potencial de los antioxidantes en tratamientos in vivo e in vitro. Rev. argent. androl. 2005; 14 (3): 90-99.
9.Adam J. Koppers, Manohar L. Garg, robert J. Aitken. Stimulation or mitochondrial reactive oxygen species production by unesterified unsaturated fatty acids in defective human spermatozoa. Free Radical Biol. 2010. Med 48 (1): 112-119.
10.Colin Williams A, Christopher Ford W. Relationship between reactive oxygen species production and lipid peroxidation in human sperm suspensions and their association with sperm function. Fertil Steril 2005; 83 (4): 929-936.
11.Moustafa M, Sharma RK, Thornton J, Mascha E, Abdel-Hafez MA, Thomas AJ, et al. Relationship between ROS production, apoptosis and DNA denaturizing in spermatozoa from patients examined for infertility. Hum Reprod 2004; 19:129-138.
12.De Jonge C. Biological basis for human capacitation. Hum Reprod 2005; 11: 205-214.
13.Zaniboni L, Cerolini S. Liquid storage of turkey semen: Changes in quality parameters, lipid composition and susceptibility to induced in vitro peroxidation in control, n3-fatty acid and alpha-tocopherol rich spermatozoa. An Reprod Sci 2009; 112: 51-65.
14.Herrero M, Chatterjee S, Lefiévre L, de Lamirande E, Gagnon C. Nitric oxide interacts with the cAMP pathway to modulate capacitation of human spermatozoa. Free Radic Biol Med 2000; 29: 522-536.
15.Schwartz M, Vissing J. Paternal inheritance of mitochondrial DNA. N Engl J Med 2002; 347: 576-580.
16.O’Brien J, Zini A. Sperm DNA integrity and male infertility. Urology 2005; 65: 16-22.
17.O’Flaherty C, Beorelgui B, Beconi MT. Participation of superoxide anion in the capacitation of cryopreserved bovine sperm. Int J Androl 2003; 26: 109-114.
18.Martín Plágaro C. Generación de mediadores lipídicos inducida por Pert- butilhidroperóxido en hepatocitos de rata [Tesis doctoral] Leioa : Universidad del País Vasco; 2001.
19.Villegas J, Schulz M, Soto L, Iglesias T, Miska W, Sánchez R. Influence of reactive oxygen species produced by activated leukocytes at the level of apoptosis in mature human spermatozoa. Fertil Steril 2005; 83: 808-810.
20.Ford WCL. Reactive oxygen species and sperm. Hum Fert 2001; 4: 77-78.
21.Oliva R. Protamines and male infertility. Hum Reprod Upd 2006; 12: 417-435.
22.Pérez Gastell P L, Pérez de Alejo J L. Métodos para medir el daño oxidativo. Rev. cuba. med. mil. 2000; 29 (3): 192-198.
23.Calamera J C, , Doncel G F, Olmedo S B, Kolm P, Acosta A A. Modified sperm stress test (MOST): a simple assay that predicts sperm-related abnormal in vitro fertilization. Hum. Reprod 1998; 13 (9): 2484-2488.
24.Bouvet B, Gatti VN, Paparella CP, Brufman AS, Solis EA, Feldman RN. Estrés espermático modificado. Respuesta inmune y funcionalidad de la membrana del espermatozoide humano. Arch Esp de Urol 2004; 57 (5): 533-537.
25.Bouvet B, Paparella CP, Feldman RN. Estrés oxidativo y su efecto sobre calidad seminal. Rev haban cienc méd. 2007; 6 (4): 1-7.
26.Calamera J C, Calamera P. El laboratorio de Andrología como ayuda diagnóstica. CEGYR. Rev Med Reprod 2003; 6 (1): 16-26.
27.Wykes S M, Krawetz S A. The structural organization of sperm chromatin. J. Biol. Chem 2003; 278 (32): 29471-29477.
28.Buffone M, Calamera J C. Evaluación de la estructura e integridad de la cromatina espermática humana en Andrología. Rev. argent. androl 2004; 13 (1): 2-14.
29.Erenpreiss J, Spano M, Erenpreisa J, Bungum M, Giwercman A. Sperm chromatin structure and male fertility: biological and clinical aspects. Asian J. Androl 2006; 8: 11-29.
30.Ollero M, Gil-Guzman E, Lopez MC, Sharma RK, Agarwal A, Larson K, Evenson D, et al. Characterization of subsets of human spermatozoa at different stages of maturation: implications in the diagnosis and treatment of male infertility. Hum Reprod 2001; 16 (9): 1912-1921.
31.Suganuma R, Yanagimachi R, Meistrich ML. Decline in fertility of Mouse sperm with abnormal chromatin during epididymal passage as revealed by ICSI. Hum Reprod 2005; 20: 3101-3108.
32.Ílvarez J G. Utilidad clínica del estudio de fragmentación del ADN espermático. Rev. argent. androl 2009; 18 (1): 3-13.
33.Jha K N, Salicioni AM, Arcelay E, Chertihin O, Kumari S, Herr JC, et al. Evidence for the involvement of proline-directed serine/threonine phosphorylation in sperm capacitation. Mol Hum Reprod 2006; 12: 781-789.
34.Smith R, Kaune H, Parodi D, Madariaga M, Morales I, Ríos R, et al. Aumento del daño en el ADN y estrés oxidativo en espermatozoides de pacientes con oligozoospermia idiopática y antecedentes de criptorquidismo. Rev Méd Chil 2007; 135 (3): 279-286.
35.Caroppo E, Niederberger C, Elhanbly S, Schoor R, Ross L, D’Amato G. Effect of cryptorchidism and retractile testes on male factor infertility: a multicenter, retrospective, chart review. Fertil Steril 2005; 83: 1581-1584.
36.Smith R, Kaune H, Parodi D, Madariaga M, Rios R, Morales I, et al. Increased sperm DNA damage in patients with varicocele: relationship with seminal oxidative stress. Hum. Reprod. 2005; 16:35-41.
37.Allamenen S, Naughton CK, Sharma RK, Thomas AJ Jr, Agarwal A. Increased seminal reactive ogygen species levels in patients with varicoceles correlate with varicocele, grade but not with testis size. Fertil Steril. 2004; 82(6):1684-1686.
38.Solis E, Gatti, AS, Brufman B, bouvet R, Conforti C, Papparella C, et al. Inmunología y deterioro de parámetros seminales en varicocele. Arch Esp. Urol. 2001; 54(8): 797-800.
39.Bustos-Obregón E, Hartley R. Ecotoxicología y daño testicular: contaminación química ambiental. Int. J. Morphol. 2008; 26(4): 833-840.
40.Perry M J. Effect of environmental and occupational pesticide exposure on human sperm: a systematic review. Hum Reprod. Update 2008; 14 (3): 322-242.
41.De La Parra I, Oizerovich S, Escobar ME. Alteraciones del eje reproductivo por enfermedades crónicas o sistémicas, sustancias tóxicas y drogas ilícitas. Rev. Reprod Hum. 2002; 2 (3):21.
42.Paparella C, Pavesi A, Feldman N, Bouvet, B. Efecto de los agroquímicos en la espermatogénesis. Rev haban cienc méd; 2011, 10(2): 1-7.
43.Bouvet B, Paparella C, Feldman R. Alteration of the sperm function associated with occupational exposition to solvents [abstract]. Abstracts from the VIII Congress-XXVI Annual Meeting December, 5-7, 2006 Sede de Gobierno de la Universidad Nacional de Rosario. Biocell 2007; 31 (1): 141.
44.Ramadan A, Agarwal A, Sharma RK, Nelson DR, Thomas AJ Jr. Effect of cigarette smoking on levels of seminal oxidative stress in infertile men: a prospective study. Fertil Steril 2002; 78: 491.
45.Bouvet B R, Paparella C, Feldman N. Efecto del tabaquismo sobre la espermatogénesis en hombres con infertilidad idiopática. Arch Esp. Urol. 2007; 60(3): 273-277.
46.Eskenazi B, Kidd SA, Marks AR, Sloter E, Block G, Wyrobek AJ. Antioxidant intake is associated with semen quality in healthy men. Hum Reprod 2005; 20: 1006-1012.
47.Keskes-Ammar L, Feki-Chakroun N, Rebai T, Sahnoun Z, Ghozzi H, Hammami S, et al. Sperm oxidative stress and the effect of an oral vitamin E and selenium supplement on semen quality in infertile men. Ach Androl 2003; 49: 83-94.
48.Lenzi A, Lombardo F, Sgrò P, Salacone P, Caponecchia L, Dondero F, et al. Use of carnitine therapy in selected casesof male factor infertility: a double-blind crossover trial Fertil Steril 2003; 79: 292-300.
49.Aitken RJ1, Smith TB, Jobling MS, Baker MA, De Iuliis GN. Oxidative stress and male reproductive health. Asian J. Androl 2014 16(1) : 31-38.
50.Bouvet B. al. Estrés oxidativo y su efecto sobre la capacidad reproductiva del espermatozoide humano. Bioquím Patol Clín 2004; 68 (1): 41.
51.Xia Wang M D, Sharma RK, Sikka SC, Thomas AJ Jr, Falcone T, Agarwal A. Oxidative stress is associated with increased apoptosis leading to spermatozoa DNA damage in patients with male factor infertility. Fertil and Steril 2003; 80 (3): 531-535.
52.Rosario Tapia Serrano. Una visión actual de la infertilidad masculina. Rev Mex Reprod 2012; 4(3):103-109.
53.Ortiz A. Factor masculino y stress oxidativo. Actual Obst y Ginecol, 2011; 1-8.


{kind=link}