










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Links relacionados
Compartilhar

 Permalink
PermalinkRevista Médica del Uruguay
versão On-line ISSN 1688-0390
Rev. Méd. Urug. vol.25 no.3 Montevideo set. 2009
Trasplante de células miogénicas: medicina regenerativa en patologías del músculo esquelético y cardíaco
Daniel Skuk, MD.*
Unidad de Investigación en Genética Humana, Centro de Investigación del Centro Hospitalario de la Universidad Laval, Quebec, Canadá
Resumen
El trasplante de células con capacidad miogénica es una estrategia experimental de medicina regenerativa. Explorada en sus comienzos como una herramienta terapéutica en el manejo de miopatías, su extrapolación al tratamiento potencial del infarto de miocardio cobra hoy mayor expansión. El presente trabajo, basado en la bibliografía y en la experiencia del autor durante más de una década dedicada a la investigación clínica y preclínica en ese terreno, pretende realizar una breve puesta al día de un tema que puede cobrar gran importancia en la medicina del presente siglo.
Palabras clave: Trasplante de Células.
Mioblastos - trasplante.
Infarto del Miocardio - terapia.
Keywords: Cell Transplantation.
Myoblasts – transplantation.
Myocardial Infarction - therapy.
* Profesor Asociado al Departamento de Anatomía y Fisiología de la Facultad de Medicina de la Universidad Laval, Quebec, Canadá.
Correspondencia: Daniel Skuk, MD.
Unité de recherche en Génétique humaine (RC9300)
Centre de recherche du Centre hospitalier de l’Université Laval
2705, boulevard Laurier, Québec, QC, G1V4G2, Canada
Correo electrónico: Daniel.Skuk@anm.ulaval.ca
Recibido: 13/4/09.
Aceptado: 20/7/09.
Introducción
El trasplante de células miogénicas implica el implante en un organismo de células capaces de formar músculo estriado. Es una estrategia en fase experimental destinada en su mayor parte al tratamiento de patologías del músculo esquelético y cardíaco. Los estudios más numerosos realizados a nivel preclínico y clínico se han focalizado esencialmente en el tratamiento de dos entidades, y en ellas se centrará el presente trabajo: las distrofias musculares (fundamentalmente las recesivas o ligadas al cromosoma X)(1,2) y el infarto de miocardio(3). Otras afecciones podrían agregarse a las anteriores, si bien hasta ahora han merecido pocos estudios(4). Sin embargo, corresponde destacar la incontinencia urinaria por deficiencias del esfínter externo, tanto por su frecuencia como por el hecho de que ha merecido ya algún ensayo clínico(5).
El uso del trasplante de células miogénicas en medicina fue propuesto en 1978 por Partridge, Grounds y Sloper, quienes sugirieron que "en pacientes portadores de miopatías hereditarias recesivas, la función muscular podría ser restaurada si se lograra que mioblastos normales fusionaran con las fibras musculares patológicas"(6). Estos autores se referían al implante de mioblastos, las células mononucleadas precursoras del músculo esquelético.
Trasplante de células en el músculo esquelético
Las propiedades potencialmente útiles del trasplante celular para el tratamiento de patologías del músculo esquelético son tres: inducir la síntesis de proteínas terapéuticas en las fibras musculares, regenerar tejido contráctil, y reponer la población de células madre.
1) Síntesis de proteínas terapéuticas. Al integrarse las células trasplantadas a las fibras musculares del receptor, la coexistencia de núcleos del donante y del receptor en un mismo sincicio determina que se sinteticen allí proteínas de ambos orígenes. Por este mecanismo, estas fibras musculares "híbridas" pueden producir la proteína cuyo déficit genético causaba una miopatía. En la distrofia muscular de Duchenne, miopatía degenerativa de origen genético ligada al cromosoma X, los pacientes sufren una pérdida muscular progresiva debida al déficit de distrofina, una proteína asociada a la membrana celular. Nuestro equipo demostró recientemente que mediante la inyección intramuscular de mioblastos de donantes normales, utilizando la técnica desarrollada por el autor, se obtenía sistemáticamente la síntesis de la distrofina del donante en fibras musculares de estos pacientes(7-9) (figura 1 C a H). Los niveles de corrección molecular en nuestros ensayos clínicos variaron de 3% a 34,5% del tejido muscular en los sitios inyectados con mioblastos normales. Estos porcentajes dependieron esencialmente de: (a) variables técnicas en la inyección de las células; (b) la calidad de las mismas, y (c) la presencia o no de rechazo inmunológico.
2) Generación de nuevo tejido contráctil. La capacidad de los mioblastos de ratón para reconstituir el tejido muscular fue observada en casos de destrucción muscular aguda, severa e irreversible(10,11). En nuestra experiencia clínica hemos observado neoformación de fibras musculares, si bien pequeñas, en pacientes que recibieron inyecciones intramusculares de mioblastos normales(7). Esta propiedad se intenta explotar clínicamente para el tratamiento de la incontinencia urinaria por deficiencias del esfínter externo de la uretra(12). El principio de este tratamiento consiste en reforzar el esfínter mediante el autotrasplante de células miogénicas cultivadas a partir de una biopsia muscular realizada al propio paciente. Un ensayo clínico preliminar reportó mejoras de la incontinencia urinaria en cinco de ocho mujeres tratadas de este modo, una de las cuales logró continencia total(5).
3) Reposición de células madre. Estudios en ratones mostraron que los mioblastos trasplantados dan origen también a células satélite, las células madre específicas del músculo esquelético(13,14). Nuestra experiencia clínica también sugiere que la misma propiedad ocurriría en el humano(7).
Tipos celulares para el trasplante
Una célula apta para trasplante puede ser una célula diferenciada o un precursor con la capacidad para diferenciarse en aquella. En el músculo esquelético, los elementos diferenciados del parénquima no son aptos para el trasplante: las fibras musculares son largos sincicios posmitóticos que no pueden ser proliferados en cultivo, ni aislados o implantados en forma adecuada. Lo más apropiado, entonces, es utilizar una célula precursora.
Células progenitoras del músculo esquelético
El músculo esquelético es uno de los tejidos altamente diferenciados que poseen capacidad regenerativa. Ello es posible gracias a la existencia de progenitores específicos: las células satélites (figura 1 A y B). Las células satélites permanecen quiescentes en la periferia de las fibras musculares hasta que estas sufren un daño; en ese momento se activan y dividen generando mioblastos, los cuales proliferan y se fusionan para formar miotubos, elementos multinucleados alargados que aumentan de volumen hasta reconstituir las fibras musculares. Las células satélite son consideradas células madre, ya que se dividen en forma asimétrica(15,16), es decir, al dividirse generan elementos diferenciados al tiempo que mantienen el pool de células madre.
Para la medicina regenerativa, la importancia de las células satélite reside en tres hechos: (a) se obtienen de un tejido accesible por un sencillo procedimiento biópsico; (b) pueden ser aisladas fácilmente mediante métodos estándar de cultivo celular, y (c) proliferan fácilmente in vitro como mioblastos (figura 1 C y D), manteniendo un fenotipo constante y pudiendo producir millones de células miogénicas en relativamente poco tiempo.
Células de la médula ósea
Dos tipos celulares de la médula ósea han sido rotulados como células madre: las células madre hematopoyéticas y un grupo de células estromales definidas como células madre mesenquimatosas. Si bien se reportó la diferenciación de células estromales en células musculares(17,18), la experiencia del autor en primates no humanos fue negativa(1). La observación en cuanto a que células circulantes derivadas de la médula ósea fusionaban ocasionalmente con fibras musculares despertó cierta expectativa en el uso potencial de esta técnica para el tratamiento de las distrofias musculares(19,20), pero luego se confirmó en ratones(21), perros(22) y pacientes(23) con distrofia muscular, que este fenómeno era irrelevante, observándose, respectivamente, apenas 0,25%, 0% y <1% de fibras musculares con corrección del defecto molecular. La especulación sobre que el mismo era debido a células madre circulantes fue puesto en jaque por experimentos que lo explicaban meramente por la fusión ocasional de monocitos o macrófagos en las fibras musculares en regeneración(24).
Mesoangioblastos
En los últimos años, un equipo italiano publicó resultados sorprendentes mediante el uso de una nueva célula a la cual se denominó mesoangioblasto(25,26). En ratones con distrofia muscular se corrigió el defecto molecular en más de 50% de las fibras musculares luego de inyecciones intraarteriales de mesoangioblastos de ratones normales(26). En perros con distrofia muscular, las inyecciones intraarteriales de mesoangioblastos normales corrigieron el defecto molecular en 4% a 70% de las fibras musculares, dependiendo del animal y del músculo analizado, reportándose además mejoras en la motricidad(25). Algunos autores, sin embargo, cuestionan la validez de la mejoría clínica de los animales, que podría deberse a la inmunosupresión(27), y quedan por determinar las potenciales consecuencias en otros órganos de una administración sistémica de grandes cantidades de células.
Vía del trasplante celular
Tras definir una célula apropiada para el trasplante, se debe determinar la técnica de implante. El desafío es importante para el músculo esquelético, ya que constituye cerca de 40% del peso corporal. Dos vías son teóricamente posibles: local e intravascular.
Implante local
Si bien es el método más frecuente, su limitante es que las células inyectadas se integran esencialmente a las fibras musculares vecinas al sitio de inyección, fenómeno descripto por el autor en primates no humanos(28-30) y en pacientes(7,9). Ello impone una cantidad elevada de inyecciones para lograr una incorporación significativa de las células injertadas. Experimentalmente se ha intentado expandir la fusión de los mioblastos implantados mediante diversas técnicas. El uso intramuscular de sustancias miotóxicas es eficaz en roedores(31), pero no siempre reproducible en primates no humanos(28,30). Inhibir la proliferación de las células satélite del receptor mediante radiación ionizante favorece la participación de las células trasplantadas en ratones(31,32). Estrategias de terapia génica ex vivo, como la modificación genética de los mioblastos para inducir la secreción de enzimas que degradan la matriz extracelular(33), no han logrado, en nuestras manos, mejorar los injertos en primates no humanos(34). Dado que por el momento no se ha conseguido extender la integración de las células implantadas más allá de los sitios de inyección utilizando técnicas aplicables en la clínica con un adecuado margen riesgo/beneficio, el autor ha considerado necesario mejorar técnicamente el implante celular mediante el uso de dispositivos que aceleren, faciliten y sistematicen la inyección intramuscular repetitiva de células. Con esa finalidad, hemos adaptado dispositivos manuales existentes para otros fines (reparto fraccionado de pequeñas cantidades de líquido en laboratorio)(35) y, en una segunda etapa, desarrollado dispositivos con cierto grado de automatización, diseñados específicamente para el implante celular repetitivo(36). También hemos propuesto, como estrategia de futuro, el desarrollo de dispositivos de implante robotizados, programables según un análisis imagenológico de las regiones a inyectar(37,38).
Administración intravascular
Dado el volumen del músculo esquelético y la inaccesibilidad de algunos lugares para inyecciones percutáneas, el trasplante se beneficiaría de una distribución intravascular. Las únicas células en las que se ha reportado una capacidad para incorporarse eficientemente a las fibras musculares a partir de una inyección intravascular son las mesoangioblastos(25,26). Hasta ahora, sin embargo, este resultado no ha sido reproducido por grupos independientes.
Supervivencia del trasplante celular
La supervivencia de las células trasplantadas deviene en el tercer factor en el éxito del injerto. Puede verse afectada por factores a corto o largo plazo.
Supervivencia a corto plazo
Dejando de lado el rechazo hiperagudo (habitualmente evitado por la detección de anticuerpos antidonante en el receptor), la supervivencia inmediata del trasplante celular puede verse perjudicada por fenómenos que no están presentes en el trasplante de órganos, como la anoikis(39), un mecanismo de apoptosis activado por la pérdida de contacto entre la célula y su sustrato. Por otro lado, si las células implantadas proliferan la mortalidad puede compensarse(40). Existe consenso en cuanto a que en el trasplante de precursores musculares un porcentaje apreciable de las células implantadas muere en los primeros tres días. En nuestras experiencias en ratones, esa muerte celular, evaluada con un marcador radioactivo, varió de 70% en el trasplante de células de cultivos primarios a 97% en el trasplante de células clonadas y modificadas genéticamente(40). El autor fue el primero en mostrar evidencias de apoptosis y necrosis en las células injertadas en el período inmediato postrasplante(40) (figura 2). Este fenómeno no compromete el injerto debido a que no todas las células mueren y a que la proliferación compensaría la mortalidad en forma parcial(41) o total(40).
Este proceso de muerte precoz no está aún bien caracterizado y existen controversias sobre sus causas. Algunas observaciones en ratones llevaron a que nuestro grupo postulara como causa la reacción inflamatoria(42), aunque un estudio posterior parecería refutar esta hipótesis(43), sugiriéndose incluso que los macrófagos podrían tener un efecto protector en los mioblastos trasplantados(44). Recientemente, el autor demostró la importancia de la isquemia en las acumulaciones intratisulares que forman células implantadas(45). Sólo una faja periférica de 100 a 200 µm se beneficia de la difusión de oxígeno y nutrientes a partir del tejido receptor, mientras que las células situadas en el centro mueren (figura 3). Este fenómeno limita el número de células que pueden implantarse por inyección directa en un único sitio: los implantes de 1 x 106, 3 x 106, 10 x 106 y 20 x 106 de células presentaron, respectivamente una media de 2%, 9%, 41% y 59% de necrosis isquémica(45).
Supervivencia a largo plazo
El único factor que parece comprometer la supervivencia a largo plazo del injerto celular es el rechazo agudo, presente en condiciones alogénicas. El rechazo agudo de las fibras musculares híbridas en condiciones de inmunosupresión inadecuada fue descripto por el autor en primates no humanos(28,29) y en pacientes que recibieron alotrasplantes de mioblastos(7) (figura 4). Los medios actuales o potenciales para evitar el rechazo agudo son la inmunosupresión, el desarrollo de tolerancia inmunológica, y el autotrasplante.
Inmunosupresión. La elección de protocolos de inmunosupresión debe ser cuidadosa, pues algunos fármacos pueden tener efectos nocivos sobre las células implantadas. Un ejemplo es la ciclofosfamida, usada en un ensayo clínico preliminar de trasplante de mioblastos(46), la cual eliminaría las células trasplantadas en la medida en que estas proliferen(47). La ciclosporina, si bien fue usada en ratones(10,48), posee propiedades que pueden afectar el éxito del injerto: inhibe la fusión de mioblastos in vitro(49), interfiere con su diferenciación(50) y puede inducir su apoptosis(50). El micofenolato mofetilo inhibe la fusión in vitro de los mioblastos de ratón(51), aunque no parece afectar el resultado del injerto en macacos(29). Los mejores aloinjertos de mioblastos en el ratón fueron obtenidos por nuestro grupo usando tacrolimus(31). Esto llevó a que lo probáramos con éxito en macacos(28-30) y que se convirtiera en nuestro inmunosupresor de elección para ensayos clínicos(7-9).
Tolerancia inmunológica. Eludir el rechazo preservando la inmunidad contra patógenos y tumores, así como evitar los efectos adversos de los inmunosupresores, es un objetivo primordial de la investigación en trasplantes. Aunque el trasplante de células miogénicas presentaría problemas específicos para el desarrollo de tolerancia inmunológica periférica o central(52), podría ofrecer otras soluciones si fuera posible manipular las células ex vivo para reducir su inmunogenicidad(53). Un estudio en el cual se modificaron genéticamente mioblastos para que expresaran HLA-G (molécula que interviene en la tolerancia materna hacia el feto) mostró que estos eran capaces de resistir a la lisis por linfocitos activados(54).
Autotrasplante. Una forma de evitar la histoincompatibilidad es el uso de células del propio paciente, manipuladas in vitro para corregir el defecto genético. El principio fue probado en ratones por nuestro equipo y otros, trasplantando mioblastos de ratones o pacientes con distrofia muscular, previamente corregidos ex vivo(55,56). También hemos demostrado la viabilidad de la estrategia en macacos(57), importante para la extrapolación al hombre. Esta táctica, sin embargo, deja la posibilidad de un rechazo a largo plazo por incompatibilidad de antígenos menores (el producto del transgen incorporado en las células)(58).

Figura 2
Figura 4
Tumorigenicidad del trasplante celular
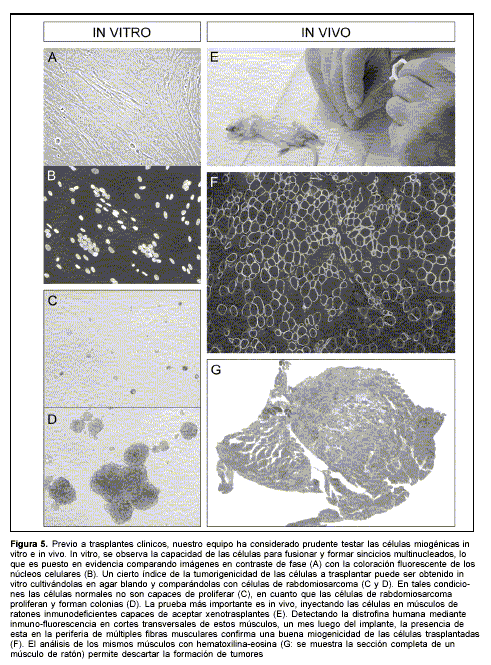
Un riesgo potencial del trasplante de células con capacidad proliferativa es la formación de neoplasmas. Este riesgo es mayor en el caso de cultivos que puedan dar origen a líneas celulares continuas o en células donde una manipulación genética pueda activar oncogenes. En el ratón, por ejemplo, se observaron rabdomiosarcomas tras el trasplante de líneas continuas de células miogénicas(10,59). Si bien no se ha reportado formación de tumores tras el trasplante de mioblastos obtenidos de cultivos primarios, nuestro equipo considera prudente excluir la tumorigenicidad de las células a trasplantar en pacientes, observando su comportamiento in vivo. Las células son trasplantadas en músculos de ratones inmunodeficientes(60), evaluándose por histopatología si hubo formación de tumores (figura 5).
Trasplante de células en el músculo cardíaco
El músculo cardíaco, al contrario del esquelético, no dispone de una célula progenitora eficaz. La capacidad proliferativa de los cardiomiocitos es motivo de controversia(61,62), y si bien podrían existir ciertos progenitores(63-65), la regeneración del miocardio es en los hechos nula. Por ello, el infarto de miocardio produce una cicatriz cuyas consecuencias son la remodelación y la insuficiencia cardíaca. Un tratamiento ideal sería sustituir la cicatriz por un tejido capaz de participar en la función de bomba del ventrículo; con ese objetivo, la idea del trasplante de células miogénicas no tardó en ser extrapolada al músculo cardíaco.
Los principales aspectos del trasplante de células miogénicas ya han sido analizados, por lo que se abordarán a continuación cuestiones específicas al músculo cardíaco.
Tipos celulares para el trasplante
Varios tipos celulares han sido utilizados en experimentos animales y en ensayos clínicos.
Cardiomiocitos
La primera célula a considerar en medicina regenerativa es aquella que forma el tejido que desea reconstruirse. Si fuera posible extraer una biopsia de miocardio del propio paciente y proliferar sus cardiomiocitos en cultivo produciendo un número adecuado para el trasplante, la ingeniería tisular del miocardio se vería facilitada. Pero los cardiomiocitos del adulto no proliferan in vitro(66) y sólo la creación de ratones transgénicos en los cuales estas células proliferaban gracias a la expresión del antígeno T del papovirus SV-40(67), permitió estudios pioneros de trasplante de cardiomiocitos(68). Los cardiomiocitos fetales, por su lado, sí proliferan in vitro, y trasplantados en el corazón de ratones y perros son capaces de formar discos intercalares con los cardiomiocitos del receptor(69), integrándose funcionalmente al miocardio(70). En ratas, el trasplante de cardiomiocitos fetales en zonas de infarto fue capaz de neoformar tejido cardíaco, limitar la expansión de la cicatriz y mejorar la función cardíaca(71). Este implante evitaba la remodelación cardíaca pero no la revertía(72).
Mioblastos
La facilidad para obtener y producir estas células, la posibilidad del trasplante autólogo, y el precedente de seguridad sentado por los trasplantes clínicos en distrofias musculares facilitaron su uso experimental en el corazón.
Estudios preclínicos. La técnica fue propuesta en 1989 por Kao y colaboradores(73). En sus primeros estudios, este equipo interpretó que los mioblastos injertados se transdiferenciaban en cardiomiocitos(73,74), lo que fue refutado luego por estudios más rigurosos(75). De hecho, las descripciones de "transdiferenciación" pueden explicarse por artefactos técnicos o por la fusión de mioblastos con cardiomiocitos del receptor(76). Taylor y colaboradores fueron los primeros en reportar una mejora de la función cardíaca tras el trasplante de mioblastos en una región necrótica del miocardio(77). Esta mejora no era producida por la neoformación de un tejido capaz de integrarse funcionalmente al miocardio, pues ambos quedaban aislados por la cicatriz. La ausencia de interacción electro-mecánica entre el tejido neoformado y el miocardio fue confirmada luego por registros intracelulares(78). Como corolario de varios estudios(79-83), se concluyó que el trasplante de mioblastos en el infarto era capaz de mejorar parcialmente la función miocárdica en modelos animales (sobre todo en roedores), aunque no por neoformación de un tejido capaz de contribuir a la contracción cardíaca, sino por cambios en las propiedades físicas de la cicatriz.
Ensayos clínicos de fase I. En el año 2000, el equipo del Dr. Philippe Menaché realizó en Francia la primera inyección intracardíaca de mioblastos autólogos en un paciente con insuficiencia cardíaca severa causada por un infarto de miocardio(84), iniciando el primer ensayo clínico de fase I(85). El mismo incluyó pacientes con cicatriz no viable y con indicación de bypass coronario (la cirugía permitía el abordaje del corazón para las inyecciones de mioblastos). El estudio mostró la viabilidad y seguridad de la técnica –con la salvedad de la aparición de arritmias– reportándose mejoras sintomáticas y estabilización de la fracción de eyección. Otros ensayos clínicos se sucedieron con protocolos y resultados esencialmente similares(86-88). En algunos pacientes se obtuvieron muestras de miocardio (por deceso o por trasplante cardíaco), donde la histología mostró acumulaciones de miotubos en la cicatriz del infarto(88,89). Como estos ensayos eran concomitantes a una cirugía de bypass coronario, no pudo dilucidarse en qué medida el beneficio era atribuible sólo al bypass o incluía el injerto. Algunos ensayos intentaron la implantación por catéter(90-92). Todos los estudios confirmaron la viabilidad del procedimiento, pero sugirieron un aumento de arritmias ventriculares.
Ensayos clínicos de fase II. Para evaluar de forma más concluyente el trasplante intracardíaco de mioblastos, el equipo del Dr. Menasché organizó un estudio clínico de fase II, multicéntrico, randomizado, en doble ciego, con control placebo y dosis escaladas(93). Noventa y siete pacientes fueron divididos en tercios que, durante un bypass coronario, recibieron respectivamente inyecciones de placebo, 400 x 106 mioblastos o 800 x 106 mioblastos. El implante no mejoró la función regional o global del ventrículo izquierdo por sobre el efecto del bypass, pero se consideró prometedor que la mayor dosis de células produjera una reversión significativa de la remodelación ventricular. Esta observación fue explicada por un potencial efecto de los mioblastos en la composición de la matriz extracelular de la cicatriz del infarto, que reduciría la fibrosis. Si ese efecto benéfico sobre la remodelación fuese persistente, podría traducirse en un mejor pronóstico para los pacientes. Con respecto a la seguridad del procedimiento, la proporción de pacientes que sufrieron arritmias no fue significativamente diferente entre los tres grupos, pero hubo más episodios de arritmia en aquellos que recibieron implantes de mioblastos. El riesgo de un aumento en la incidencia de arritmias, de hecho, parecería existir en otros tipos de trasplante celular en el corazón (aunque con menos frecuencia que con los mioblastos), y las células implantadas lo producirían por cualquiera de los tres mecanismos arritmogénicos mayores: reentrada, automatismo y actividad gatillada(94).
Progenitores cardíacos
Se han detectado escasas células madre en el miocardio de ratones(95-97) y se describieron cardioblastos en el miocardio posnatal de ratones, ratas y humanos(98). Los mesoangioblastos también estarían presentes en el miocardio y podrían diferenciarse en cardiomiocitos(99). Ciertas evidencias sugieren además la presencia de células circulantes capaces de diferenciarse en miocardio. Las observaciones en roedores(63) parecen confirmarse en el hombre tras el análisis del tejido cardíaco de necropsias realizadas en varones que habían recibido un trasplante de corazón de mujer(64). La detección del cromosoma Y permitió inferir que 0,04% de los cardiomiocitos provenían del receptor y que esto ocurría en zonas de lesión. El uso de tales progenitores en medicina regenerativa dependería de la capacidad para aislarlos y proliferarlos en cantidad suficiente para el injerto, sin perder sus propiedades biológicas. Sin embargo, se debe destacar su ineficiencia para regenerar el miocardio en forma espontánea.
Células madre embrionarias
Estudios preliminares en roedores recurrieron al implante intramiocárdico de células madre embrionarias a fin de observar si ello bastaba para inducir su diferenciación en cardiomiocitos(100, 101). Los resultados fueron negativos y el implante generó teratomas, descartando el implante de células madre embrionarias indiferenciadas. Como la opción más lógica sería diferenciar las células madre embrionarias in vitro previo al trasplante, las condiciones de cultivo para ello comienzan a estar más definidas(102). Experiencias en ratones inmunodeficientes mostraron la capacidad de los cardiomiocitos derivados de células madre embrionarias humanas para regenerar el miocardio y mejorar la función cardíaca en zonas de infarto(103), aunque esa mejora no se mantenía a largo plazo(104). Aun dejando de lado los obstáculos ético-religiosos, ciertos detalles técnicos deben ser resueltos antes de una utilización clínica de cardiomiocitos derivados de células madre embrionarias; esencialmente, una separación específica, segura y con alto rendimiento. Por lo demás, fuera de un contexto de clonación terapéutica, su uso implica condiciones alogénicas y el recurso a inmunosupresión. Algunos autores pretenden que las células madre embrionarias tendrían privilegio inmunológico, es decir, serían "invisibles" al sistema inmunitario en un contexto alogénico e incluso xenogénico(105,106). Esta hipótesis, sin embargo, deja de lado el hecho de que estas células deben diferenciarse y de que las propiedades antigénicas de los cardiomiocitos obtenidos de células madre embrionarias no parecen diferir de sus homólogos adultos(107). Una vía interesante para la obtención autóloga de células pluripotentes con propiedades similares a las células madre embrionarias, eludiendo las controversias éticas, sería la reprogramación in vitro de células somáticas como los fibroblastos(108,109), posibilidad que está cobrando actualmente mucho interés(110). Se ha dado a estas células la denominación de células pluripotentes inducidas.
Células de la médula ósea
Aunque ciertos estudios sugieren que las células madre mesenquimatosas podrían diferenciarse en cardiomiocitos(111), su inyección intracardíaca podría comportar riesgos de formación ectópica de otros tejidos mesodérmicos. El implante intracardíaco de médula ósea en ratas, por ejemplo, produjo tejido óseo, atribuible al potencial osteogénico de las células madre mesenquimatosas(112). Un equipo interpretó que las células madre hematopoyéticas eran capaces de neoformar miocardio en la cicatriz de infarto en ratones(113), pero estudios posteriores en ratones(114), ovinos(115) y primates no humanos(116), fueron negativos, y se comprobó que las células madre hematopoyéticas inyectadas en el corazón se diferencian sólo en células sanguíneas(117).
Siendo el trasplante de médula ósea una técnica estándar en hematología, el paso de los experimentos en animales a ensayos clínicos fue rápido, usándose en forma autóloga tanto médula ósea no fraccionada como poblaciones seleccionadas (CD34+ y CD133+), administradas por vía intracoronaria e intramiocárdica(118). Si bien algunos estudios reportaron mejoras leves de la función cardíaca, pocos fueron randomizados y controlados y los resultados son contradictorios. Por otro lado, la inyección de células CD133+ por vía intracoronaria podría causar efectos adversos en las arterias coronarias(119,120).
Causas de la mejora postrasplante (figura 6)
El propósito original del trasplante celular intracardíaco era la regeneración de un tejido contráctil capaz de interactuar electro-mecánicamente con el tejido cardíaco del receptor. Pese a ciertos datos al inicio contradictorios, es evidente que los trasplantes de mioblastos y de células de la médula ósea no alcanzaron ese objetivo. Las mejoras de la función cardíaca observadas en los modelos animales y en los ensayos clínicos deben explicarse por otros mecanismos. La hipótesis predominante es que los injertos celulares limitan la remodelación del ventrículo izquierdo. Se ha propuesto que las acumulaciones de células trasplantadas engrosarían la pared del infarto y limitarían la dilatación del ventrículo(121). También se ha sugerido un efecto parácrino del implante, por el cual la matriz extracelular ganaría en propiedades elásticas(122). Más allá de dilucidar los mecanismos de los beneficios reportados, lo cierto es que estos últimos han sido muy limitados y el objetivo principal de la medicina regenerativa en este campo sigue siendo el señalado al inicio del párrafo.
Conclusiones
El progreso en temáticas relacionadas a la medicina regenerativa –trasplantes celulares, ingeniería tisular y terapia génica– permite cierto optimismo en prever, para un futuro cercano, la posibilidad de ofrecer mejores tratamientos para diferentes patologías del músculo esquelético y cardíaco. Los ensayos clínicos han permitido verificar la viabilidad y seguridad de algunos métodos, y aunque los resultados clínicos han sido modestos o marginales, los caminos a seguir parecen presentarse con más claridad, como se resume a continuación.

Figura 6
En el músculo esquelético, de confirmarse la capacidad de los mesoangioblastos para corregir el defecto molecular en un volumen significativo de músculo tras una inyección intravascular, sin producir efectos secundarios en otros órganos, su uso podría ser clave en el tratamiento de las miopatías genéticas recesivas o ligadas al cromosoma X en las cuales existe una necrosis constante de las fibras musculares (elemento necesario para la integración eficaz de las células implantadas por vía sistémica). En otras miopatías genéticas sin necrosis prominente, así como en los casos de degeneración muscular avanzada con sustitución del parénquima contráctil por fibrosis o tejido adiposo, el implante intramuscular de precursores musculares como los mioblastos nos sigue pareciendo la única estrategia adecuada. Lo mismo se aplicaría a déficits musculares localizados, como el caso del esfínter externo de la uretra en la incontinencia urinaria. Quedan por refinar las estrategias de implante que aseguren una integración óptima de las células inyectadas en el tejido receptor, así como los métodos que aseguren una adecuada neoformación de tejido muscular cuando ello es necesario. Para el implante intramuscular en grandes regiones, creemos necesaria la automatización en mayor o menor grado de la inyección de células.
El tratamiento efectivo del infarto de miocardio necesitará el implante de células cuyo efecto no se limite a mejorar en forma restringida las características físicas de la cicatriz del infarto. Un tratamiento efectivo podrá ser posible sólo mediante el implante intracardíaco de células capaces de desarrollar el programa de diferenciación cardiogénica, es decir, de diferenciarse en un fenotipo de cardiomiocito o equivalente que permita la propagación eléctrica, el acoplamiento electromecánico y la contracción. Las células madre embrionarias se presentan al día de hoy como las más hábiles para dar lugar eficientemente a tal diferenciación (previo al trasplante), pero los problemas éticos e inmunológicos ligados a su utilización han puesto en la mira de los investigadores el desarrollo de las células pluripotentes inducidas. En todo caso, será necesario estandarizar cuantitativamente los parámetros del trasplante, especialmente en lo que concierne a la cantidad de células por inyección y a la topografía y densidad de las mismas, de forma de asegurar una regeneración óptima y reproducible del tejido cardíaco.
Creemos del caso finalizar con un llamado de atención. Dado el creciente y a veces confuso cúmulo de estudios en medicina regenerativa (no siempre rigurosos y en algunos casos dudosos) es importante un abordaje cauto y crítico del tema: muchas novedades "prometedoras" acaban en lo irrepetible o lo refutado. Esta cautela es especialmente pertinente para evitar precipitados usos clínicos. Las aplicaciones médicas requieren una estandardización de la producción de las células adecuadas así como de las técnicas de trasplante, y ello debe realizarse en modelos animales apropiados, definiéndose las variables que permitan obtener resultados claros y reproducibles.
Summary
Miogenic cells transplantions is an experimental technique in regenerative medicine. Intitially explored as a therapeutic tool in the treatment of myopathies, its extrapolation for the potential treatment of myocardial infarction currently spreads its scope of application. This study, based on bibliography and the author’s experience during over a decade of clinical and preclinical research in the field, attempts to provide an update in a highly important topic for this century’s medicine.
Résumé
La greffe de cellules à capacité myogénique est une stratégie expérimentale de médecine régénérative. Considérée au début comme un outil thérapeutique dans le traitement des myopathies, son usage au traitement potentiel de l’infarctus du myocarde est plus large de nos jours. Ce rapport, basé sur la bibliographie et l’expérience de l’auteur récoltée pendant plus d’une décennie consacrée à la recherche clinique et préclinique, prétend faire une brève mise à la page d’un sujet qui est censé avoir une grande incidence sur la médecine de ce siècle.
Resumo
O transplante de células com capacidade miogênica é uma estratégia experimental da medicina regenerativa. Explorada a principio como uma ferramenta terapêutica no manejo das miopatias, sua extrapolação ao tratamento potencial do infarto de miocárdio possibilitou uma maior expansão. Este trabalho, baseado na bibliografia e na experiência do autor durante mais de uma década dedicada à pesquisa clínica e pré-clínica nessa área, pretende realizar una breve atualização de um tema que pode ter grande importância na medicina deste século.
Bibliografía
1. Skuk D. Myoblast transplantation for inherited myopathies: a clinical approach. Expert Opin Biol Ther 2004; 4(12): 1871-85.
2. Skuk D, Tremblay JP. Cell therapies for inherited myopathies. Curr Opin Rheumatol 2003; 15(6): 723-9.
3. Vilquin JT, Marolleau JP, Hagege A, Menasche P, Fiszman M, Schwartz K. Cell transplantation for post-ischemic heart failure. Arch Mal Coeur Vaiss 2002; 95(12): 1219-25.
4. Skuk D, Tremblay JP. Progress in myoblast transplantation: a potential treatment of dystrophies. Microsc Res Tech 2000; 48(3-4): 213-22.
5. Carr LK, Steele D, Steele S, Wagner D, Pruchnic R, Jankowski R, et al. 1-year follow-up of autologous muscle-derived stem cell injection pilot study to treat stress urinary incontinence. Int Urogynecol J Pelvic Floor Dysfunct 2008; 19(6): 881-3.
6. Partridge TA, Grounds M, Sloper JC. Evidence of fusion between host and donor myoblasts in skeletal muscle grafts. Nature 1978; 273(5660): 306-8.
7. Skuk D, Goulet M, Roy B, Chapdelaine P, Bouchard JP, Roy R, et al. Dystrophin expression in muscles of Duchenne muscular dystrophy patients after high-density injections of normal myogenic cells. J Neuropathol Exp Neurol 2006; 65(4): 371-86.
8. Skuk D, Goulet M, Roy B, Piette V, Côté CH, Chapdelaine P, et al. First test of a "high-density injection" protocol for myogenic cell transplantation throughout large volumes of muscles in a Duchenne muscular dystrophy patient: eighteen months follow-up. Neuromuscul Disord 2007; 17(1): 38-46.
9. Skuk D, Roy B, Goulet M, Chapdelaine P, Bouchard JP, Roy R, et al. Dystrophin expression in myofibers of Duchenne muscular dystrophy patients following intramuscular injections of normal myogenic cells. Mol Ther 2004; 9(3): 475-82.
10. Wernig A, Irintchev A, Lange G. Functional effects of myoblast implantation into histoincompatible mice with or without immunosuppression. J Physiol 1995; 484(Pt 2): 493-504.
11. Alameddine HS, Louboutin JP, Dehaupas M, Sébille A, Fardeau M. Functional recovery induced by satellite cell grafts in irreversibly injured muscles. Cell Transplant 1994; 3(1): 3-14.
12. Yokoyama T, Huard J, Chancellor MB. Myoblast therapy for stress urinary incontinence and bladder dysfunction. World J Urol 2000; 18(1): 56-61.
13. Heslop L, Beauchamp JR, Tajbakhsh S, Buckingham ME, Partridge TA, Zammit PS. Transplanted primary neonatal myoblasts can give rise to functional satellite cells as identified using the Myf5(nlacZl+) mouse. Gene Ther 2001; 8(10): 778-83.
14. Ehrhardt J, Brimah K, Adkin C, Partridge T, Morgan J. Human muscle precursor cells give rise to functional satellite cells in vivo. Neuromuscul Disord 2007; 17(8): 631-8.
15. Shinin V, Gayraud-Morel B, Gomes D, Tajbakhsh S. Asymmetric division and cosegregation of template DNA strands in adult muscle satellite cells. Nat Cell Biol 2006; 8(7): 677-87.
16. Collins CA, Olsen I, Zammit PS, Heslop L, Petrie A, Partridge TA, et al. Stem cell function, self-renewal, and behavioral heterogeneity of cells from the adult muscle satellite cell niche. Cell 2005; 122(2): 289-301.
17. Grigoriadis AE, Heersche JN, Aubin JE. Differentiation of muscle, fat, cartilage, and bone from progenitor cells present in a bone-derived clonal cell population: effect of dexamethasone. J Cell Biol 1988; 106(6): 2139-51.
18. Wakitani S, Saito T, Caplan AI. Myogenic cells derived from rat bone marrow mesenchymal stem cells exposed to 5-azacytidine. Muscle Nerve 1995; 18(12): 1417-26.
19. Ferrari G, Cusella-De Angelis G, Coletta M, Paolucci E, Stornaiuolo A, Cossu G, et al. Muscle regeneration by bone marrow-derived myogenic progenitors. Science 1998; 279(5356): 1528-30.
20. Gussoni E, Soneoka Y, Strickland CD, Buzney EA, Khan MK, Flint AF, et al. Dystrophin expression in the mdx mouse restored by stem cell transplantation. Nature 1999; 401(6751): 390-4.
21. Ferrari G, Stornaiuolo A, Mavilio F. Failure to correct murine muscular dystrophy. Nature 2001; 411(6841):1014-5.
22. Dell’Agnola C, Wang Z, Storb R, Tapscott SJ, Kuhr CS, Hauschka SD, et al. Hematopoietic stem cell transplantation does not restore dystrophin expression in Duchenne muscular dystrophy dogs. Blood 2004; 104(13): 4311-8.
23. Gussoni E, Bennett RR, Muskiewicz KR, Meyerrose T, Nolta JA, Gilgoff I, et al. Long-term persistence of donor nuclei in a Duchenne muscular dystrophy patient receiving bone marrow transplantation. J Clin Invest 2002; 110(6): 807-14.
24. Camargo FD, Green R, Capetenaki Y, Jackson KA, Goodell MA. Single hematopoietic stem cells generate skeletal muscle through myeloid intermediates. Nat Med 2003; 9(12): 1520-7.
25. Sampaolesi M, Blot S, D’Antona G, Granger N, Tonlorenzi R, Innocenzi A, et al. Mesoangioblast stem cells ameliorate muscle function in dystrophic dogs. Nature 2006; 444(7119): 574-9.
26. Sampaolesi M, Torrente Y, Innocenzi A, Tonlorenzi R, D’Antona G, Pellegrino MA, et al. Cell therapy of alpha-sarcoglycan null dystrophic mice through intra-arterial delivery of mesoangioblasts. Science 2003; 301(5632): 487-92.
27. Grounds MD, Davies KE. The allure of stem cell therapy for muscular dystrophy. Neuromuscul Disord 2007; 17(3): 206-8.
28. Skuk D, Roy B, Goulet M, Tremblay JP. Successful myoblast transplantation in primates depends on appropriate cell delivery and induction of regeneration in the host muscle. Exp Neurol 1999; 155(1): 22-30.
29. Skuk D, Goulet M, Roy B, Tremblay JP. Efficacy of myoblast transplantation in nonhuman primates following simple intramuscular cell injections: toward defining strategies applicable to humans. Exp Neurol 2002; 175(1): 112-26.
30. Skuk D, Goulet M, Roy B, Tremblay JP. Myoblast transplantation in whole muscle of nonhuman primates. J Neuropathol Exp Neurol 2000; 59(3): 197-206.
31. Kinoshita I, Vilquin JT, Guérette B, Asselin I, Roy R, Tremblay JP. Very efficient myoblast allotransplantation in mice under FK506 immunosuppression. Muscle Nerve 1994; 17(12): 1407-15.
32. Morgan JE, Hoffman EP, Partridge TA. Normal myogenic cells from newborn mice restore normal histology to degenerating muscles of the mdx mouse. J Cell Biol 1990; 111(6 Pt 1): 2437-49.
33. El Fahime E, Mills P, Lafreniere JF, Torrente Y, Tremblay JP. The urokinase plasminogen activator: an interesting way to improve myoblast migration following their transplantation. Exp Cell Res 2002; 280(2): 169-78.
34. Lafreniere JF, Caron MC, Skuk D, Goulet M, Cheikh AR, Tremblay JP. Growth factor coinjection improves the migration potential of monkey myogenic precursors without affecting cell transplantation success. Cell Transplant 2009; 18(7):719-30.
35. Skuk D, Goulet M, Tremblay JP. Use of repeating dispensers to increase the efficiency of the intramuscular myogenic cell injection procedure. Cell Transplant 2006; 15(7): 659-63.
36. Richard PL, Gosselin C, Goulet M, Paradis M, Tremblay JP, Skuk D. A specific semi-manual device for rapid repetitive intramuscular cell implantation. Tissue Eng 2009; 15: 724.
37. Skuk D, Tremblay JP. Myoblast transplantation. In: Atala A, Lanza R. Methods of tissue engineering. San Diego: Academic Press, 2001: 1083-98.
38. Skuk D, Tremblay JP. "Engineering" myoblast transplantation. Graft 2001; 4: 558-70.
39. Zvibel I, Smets F, Soriano H. Anoikis: roadblock to cell transplantation? Cell Transplant 2002; 11(7): 621-30.
40. Skuk D, Caron NJ, Goulet M, Roy B, Tremblay JP. Resetting the problem of cell death following muscle-derived cell transplantation: detection, dynamics and mechanisms. J Neuropathol Exp Neurol 2003; 62(9): 951-67.
41. Beauchamp JR, Morgan JE, Pagel CN, Partridge TA. Dynamics of myoblast transplantation reveal a discrete minority of precursors with stem cell-like properties as the myogenic source. J Cell Biol 1999; 144(6): 1113-22.
42. Guérette B, Skuk D, Célestin F, Huard C, Tardif F, Asselin I, et al. Prevention by anti-LFA-1 of acute myoblast death following transplantation. J Immunol 1997; 159(5): 2522-31.
43. Sammels LM, Bosio E, Fragall CT, Grounds MD, Van Rooijen N, Beilharz MW. Innate inflammatory cells are not responsible for early death of donor myoblasts after myoblast transfer therapy. Transplantation 2004; 77(12): 1790-7.
44. Chazaud B, Sonnet C, Lafuste P, Bassez G, Rimaniol AC, Poron F, et al. Satellite cells attract monocytes and use macrophages as a support to escape apoptosis and enhance muscle growth. J Cell Biol 2003; 163(5): 1133-43.
45. Skuk D, Paradis M, Goulet M, Tremblay JP. Ischemic central necrosis in pockets of transplanted myoblasts in nonhuman primates: implications for cell-transplantation strategies. Transplantation 2007; 84(10): 1307-15.
46. Karpati G, Ajdukovic D, Arnold D, Gledhill RB, Guttman R, Holland P, et al. Myoblast transfer in Duchenne muscular dystrophy. Ann Neurol 1993; 34(1): 8-17.
47. Vilquin JT, Kinoshita I, Roy R, Tremblay JP. Cyclophosphamide immunosuppression does not permit successful myoblast allotransplantation in mouse. Neuromuscul Disord 1995; 5(6): 511-7.
48. Irintchev A, Zweyer M, Wernig A. Cellular and molecular reactions in mouse muscles after myoblast implantation. J Neurocytol 1995; 24(4): 319-31.
49. Hardiman O, Sklar RM, Brown RH Jr. Direct effects of cyclosporin A and cyclophosphamide on differentiation of normal human myoblasts in culture. Neurology 1993; 43(7): 1432-4.
50. Hong F, Lee J, Song JW, Lee SJ, Ahn H, Cho JJ, et al. Cyclosporin A blocks muscle differentiation by inducing oxidative stress and inhibiting the peptidyl-prolyl-cis-trans isomerase activity of cyclophilin A: cyclophilin A protects myoblasts from cyclosporin A-induced cytotoxicity. Faseb J 2002; 16(12): 1633-5.
51. Camirand G, Caron NJ, Asselin I, Tremblay JP. Combined immunosuppression of mycophenolate mofetil and FK506 for myoblast transplantation in mdx mice. Transplantation 2001; 72(1): 38-44.
52. Camirand G, Stéphan L, Rousseau J, Sackett MK, Caron NJ, Mills P, et al. Central tolerance to myogenic cell transplants does not include muscle neoantigens. Transplantation 2008; 85(12): 1791-801.
53. Gill RG, Wolf L. Immunobiology of cellular transplantation. Cell Transplant 1995; 4(4): 361-70.
54. Wiendl H, Mitsdoerffer M, Hofmeister V, Wischhusen J, Weiss EH, Dichgans J, et al. The non-classical MHC molecule HLA-G protects human muscle cells from immune-mediated lysis: implications for myoblast transplantation and gene therapy. Brain 2003; 126(Pt 1): 176-85.
55. Floyd SS Jr, Clemens PR, Ontell MR, Kochanek S, Day CS, Yang J, et al. Ex vivo gene transfer using adenovirus-mediated full-length dystrophin delivery to dystrophic muscles. Gene Ther 1998; 5(1): 19-30.
56. Moisset PA, Skuk D, Asselin I, Goulet M, Roy B, Karpati G, et al. Successful transplantation of genetically corrected DMD myoblasts following ex vivo transduction with the dystrophin minigene. Biochem Biophys Res Commun 1998; 247(1): 94-9.
57. Quenneville SP, Chapdelaine P, Skuk D, Paradis M, Goulet M, Rousseau J, et al. Autologous transplantation of muscle precursor cells modified with a lentivirus for muscular dystrophy: human cells and primate models. Mol Ther 2007; 15(2): 431-8.
58. Ohtsuka Y, Udaka K, Yamashiro Y, Yagita H, Okumura K. Dystrophin acts as a transplantation rejection antigen in dystrophin- deficient mice: implication for gene therapy. J Immunol 1998; 160(9): 4635-40.
59. Wernig A, Irintchev A, Hartling A, Stephan G, Zimmer-mann K, Starzinski-Powitz A. Formation of new muscle fibres and tumours after injection of cultured myogenic cells. J Neurocytol 1991; 20(12): 982-97.
60. Skuk D, Furling D, Bouchard JP, Goulet M, Roy B, Lacroix Y, et al. Transplantation of human myoblasts in SCID mice as a potential muscular model for myotonic dystrophy. J Neuropathol Exp Neurol 1999; 58(9): 921-31.
61. Soonpaa MH, Field LJ. Survey of studies examining mammalian cardiomyocyte DNA synthesis. Circ Res 1998; 83(1): 15-26.
62. Beltrami AP, Urbanek K, Kajstura J, Yan SM, Finato N, Bussani R, et al. Evidence that human cardiac myocytes divide after myocardial infarction. N Engl J Med 2001; 344(23): 1750-7.
63. Jackson KA, Majka SM, Wang H, Pocius J, Hartley CJ, Majesky MW, et al. Regeneration of ischemic cardiac muscle and vascular endothelium by adult stem cells. J Clin Invest 2001; 107(11): 1395-402.
64. Laflamme MA, Myerson D, Saffitz JE, Murry CE. Evidence for cardiomyocyte repopulation by extracardiac progenitors in transplanted human hearts. Circ Res 2002; 90(6): 634-40.
65. Quaini F, Urbanek K, Beltrami AP, Finato N, Beltrami CA, Nadal-Ginard B, et al. Chimerism of the transplanted heart. N Engl J Med 2002; 346(1): 5-15.
66. Claycomb WC. Cardiac muscle cell proliferation and cell differentiation in vivo and in vitro. Adv Exp Med Biol 1983; 161: 249-65.
67. Steinhelper ME, Lanson NA Jr, Dresdner KP, Delcarpio JB, Wit AL, Claycomb WC, et al. Proliferation in vivo and in culture of differentiated adult atrial cardiomyocytes from transgenic mice. Am J Physiol 1990; 259(6 Pt 2): H1826-34.
68. Koh GY, Soonpaa MH, Klug MG, Field LJ. Long-term survival of AT-1 cardiomyocyte grafts in syngeneic myocardium. Am J Physiol 1993; 264(5 Pt 2): H1727-33.
69. Koh GY, Soonpaa MH, Klug Mg, Pride HP, Cooper BJ, Zipes DP, et al. Stable fetal cardiomyocyte grafts in the hearts of dystrophic mice and dogs. J Clin Invest 1995; 96(4): 2034-42.
70. Rubart M, Pasumarthi KB, Nakajima H, Soonpaa MH, Nakajima HO, Field LJ. Physiological coupling of donor and host cardiomyocytes after cellular transplantation. Circ Res 2003; 92(11): 1217-24.
71. Li RK, Jia ZQ, Weisel RD, Mickle DA, Zhang J, Mohabeer MK, et al. Cardiomyocyte transplantation improves heart function. Ann Thorac Surg 1996; 62(3): 654-60.
72. Sakakibara Y, Tambara K, Lu F, Nishina T, Nagaya N, Nishimura K, et al. Cardiomyocyte transplantation does not reverse cardiac remodeling in rats with chronic myocardial infarction. Ann Thorac Surg 2002; 74(1): 25-30.
73. Kao RL, Rizzo C, Magovern GI. Satellite cell for myocardial regeneration. Physiologist 1989; 32: 220.
74. Kao RL. Regeneration of injured myocardium from implanted satellite cells. Circulation 1991; (Suppl. II) 84: II-386.
75. Reinecke H, Poppa V, Murry Ce. Skeletal muscle stem cells do not transdifferentiate into cardiomyocytes after cardiac grafting. J Mol Cell Cardiol 2002; 34(2): 241-9.
76. Reinecke H, Minami E, Poppa V, Murry CE. Evidence for fusion between cardiac and skeletal muscle cells. Circ Res 2004; 94(6): e56-60.
77. Taylor DA, Atkins BZ, Hungspreugs P, Jones TR, Reedy MC, Hutcheson KA, et al. Regenerating functional myocardium: improved performance after skeletal myoblast transplantation. Nat Med 1998; 4(8): 929-33.
78. Leobon B, Garcin I, Menasche P, Vilquin JT, Audinat E, Charpak S. Myoblasts transplanted into rat infarcted myocardium are functionally isolated from their host. Proc Natl Acad Sci USA 2003; 100(13): 7808-11.
79. Al Attar N, Carrion C, Ghostine S, Garcin I, Vilquin JT, Hagège AA, et al. Long-term (1 year) functional and histological results of autologous skeletal muscle cells transplantation in rat. Cardiovasc Res 2003; 58(1): 142-8.
80. Murry CE, Wiseman RW, Schwartz SM, Hauschka SD. Skeletal myoblast transplantation for repair of myocardial necrosis. J Clin Invest 1996; 98(11):2512-23.
81. Tambara K, Sakakibara Y, Sakaguchi G, Lu F, Premaratne GU, Lin X, et al. Transplanted skeletal myoblasts can fully replace the infarcted myocardium when they survive in the host in large numbers. Circulation 2003; 108(Suppl 1): II259-63.
82. Pouzet B, Vilquin JT, Hagege AA, Scorsin M, Messas E, Fiszman M, et al. Intramyocardial transplantation of autologous myoblasts: can tissue processing be optimized? Circulation 2000; 102(19 Suppl 3): III210-5.
83. Scorsin M, Hagège A, Vilquin JT, Fiszman M, Marotte F, Samuel JL, et al. Comparison of the effects of fetal cardiomyocyte and skeletal myoblast transplantation on postinfarction left ventricular function. J Thorac Cardiovasc Surg 2000; 119(6): 1169-75.
84. Menasché P, Hagège AA, Scorsin M, Pouzet B, Desnos M, Duboc D, et al. Myoblast transplantation for heart failure. Lancet 2001; 357(9252): 279-80.
85. Menasché P, Hagège AA, Vilquin JT, Desnos M, Abergel E, Pouzet B, et al. Autologous skeletal myoblast transplantation for severe postinfarction left ventricular dysfunction. J Am Coll Cardiol 2003; 41(7): 1078-83.
86. Gavira JJ, Herreros J, Pérez A, García-Velloso MJ, Barba J, Martin-Herrero F, et al. Autologous skeletal myoblast transplantation in patients with nonacute myocardial infarction: 1-year follow-up. J Thorac Cardiovasc Surg 2006; 131(4): 799-804.
87. Siminiak T, Kalawki R, Fiszer D, Jerzykowska O, Rzezniczak J, Rozwadowska N, et al. Autologous skeletal myoblast transplantation for the treatment of postinfarction myocardial injury: phase I clinical study with 12 months of follow-up. Am Heart J 2004; 148(3): 531-7.
88. Dib N, Michler RE, Pagani FD, Wright S, Kereiakes DJ, Lengerich R, et al. Safety and feasibility of autologous myoblast transplantation in patients with ischemic cardiomyopathy: four-year follow-up. Circulation 2005; 112(12): 1748-55.
89. Hagège AA, Carrion C, Menasché P, Vilquin JT, Duboc D, Marolleau JP, et al. Viability and differentiation of autologous skeletal myoblast grafts in ischaemic cardiomyopathy. Lancet 2003; 361(9356): 491-2.
90. Siminiak T, Burchardt P, Kurpisz M. Postinfarction heart failure: surgical and trans-coronary-venous transplantation of autologous myoblasts. Nat Clin Pract Cardiovasc Med 2006; 3(Suppl 1): S46-51.
91. Biagini E, Valgimigli M, Smits PC, Poldermans D, Schinkel AF, Rizzello V, et al. Stress and tissue Doppler echocardiographic evidence of effectiveness of myoblast transplantation in patients with ischaemic heart failure. Eur J Heart Fail 2006; 8(6): 641-8.
92. Ince H, Petzsch M, Rehders TC, Chatterjee T, Nienaber CA. Transcatheter transplantation of autologous skeletal myoblasts in postinfarction patients with severe left ventricular dysfunction. J Endovasc Ther 2004; 11(6): 695-704.
93. Menasché P, Alfieri O, Janssens S, McKenna W, Reichenspurner H, Trinquart L, et al. The Myoblast Autologous Grafting in Ischemic Cardiomyopathy (MAGIC) trial: first randomized placebo-controlled study of myoblast transplantation. Circulation 2008; 117(9): 1189-200.
94. Menasché P. Stem cell therapy for heart failure: are arrhythmias a real safety concern? Circulation 2009; 119(20): 2735-40.
95. Hierlihy AM, Seale P, Lobe CG, Rudnicki MA, Megeney LA. The post-natal heart contains a myocardial stem cell population. FEBS Lett 2002; 530(1-3): 239-43.
96. Beltrami AP, Barlucchi L, Torella D, Baker M, Limana F, Chimenti S, et al. Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell 2003; 114(6): 763-76.
97. Oh H, Bradfute SB, Gallardo TD, Nakamura T, Gaussin V, Mishina Y, et al. Cardiac progenitor cells from adult myocardium: homing, differentiation, and fusion after infarction Proc Natl Acad Sci USA 2003; 100(21): 12313-8.
98. Laugwitz KL, Moretti A, Lam J, Gruber P, Chen Y, Woodard S, et al. Postnatal isl1+ cardioblasts enter fully differentiated cardiomyocyte lineages. Nature 2005; 433(7026): 647-53.
99. Galvez BG, Sampaolesi M, Barbuti A, Crespi A, Covarello D, Brunelli S, et al. Cardiac mesoangioblasts are committed, self-renewable progenitors, associated with small vessels of juvenile mouse ventricle. Cell Death Differ 2008; 15(9): 1417-28.
100 Leor J, Gerecht S, Cohen S, Miller L, Holbova R, Ziskind A, et al. Human embryonic stem cell transplantation to repair the infarcted myocardium. Heart 2007; 93(10):1278-84.
101. Nussbaum J, Minami E, Laflamme MA, Virag JA, Ware CB, Masino A, et al. Transplantation of undifferentiated murine embryonic stem cells in the heart: teratoma formation and immune response. Faseb J 2007; 21(7): 1345-57.
102. Steel D, Hyllner J, Sartipy P. Cardiomyocytes derived from human embryonic stem cells - characteristics and utility for drug discovery. Curr Opin Drug Discov Devel 2009; 12(1): 133-40.
103. Laflamme MA, Chen KY, Naumova AV, Muskheli V, Fugate JA, Dupras SK, et al. Cardiomyocytes derived from human embryonic stem cells in pro-survival factors enhance function of infarcted rat hearts. Nat Biotechnol 2007; 25(9): 1015-24.
104. van Laake LW, Passier R, DoevendansPA, Mummery CL. Human embryonic stem cell-derived cardiomyocytes and cardiac repair in rodents. Circ Res 2008; 102(9): 1008-10.
105. Kehat I, Khimovich L, Caspi O, Gepstein A, Shofti R, Arbel G, et al. Electromechanical integration of cardiomyocytes derived from human embryonic stem cells. Nat Biotechnol 2004; 22(10): 1282-9.
106. Ménard C, Hagège AA, Agbulut O, Barro M, Morichetti MC, Brasselet C, et al. Transplantation of cardiac-committed mouse embryonic stem cells to infarcted sheep myocardium: a preclinical study. Lancet 2005; 366(9490): 1005-12.
107. Swijnenburg RJ, Tanaka M, Vogel H, Baker J, Kofidis T, Gunawan F, et al. Embryonic stem cell immunogenicity increases upon differentiation after transplantation into ischemic myocardium. Circulation 2005; 112(9 Suppl): I166-72.
108. Wernig M, Meissner A, Foreman R, Brambrink T, Ku M, Hochedlinger K, et al. In vitro reprogramming of fibroblasts into a pluripotent ES-cell-like state. Nature 2007; 448(7151): 318-24.
109. Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007; 131(5): 861-72.
110. Yuasa S, Fukuda K. Recent advances in cardiovascular regenerative medicine: the induced pluripotent stem cell era. Expert Rev Cardiovasc Ther 2008; 6(6): 803-10.
111. Fukuda K. Application of mesenchymal stem cells for the regeneration of cardiomyocyte and its use for cell transplantation therapy. Hum Cell 2003; 16(3): 83-94.
112. Yoon YS, Park JS, Tkebuchava T, Luedeman C, Losordo DW. Unexpected severe calcification after transplantation of bone marrow cells in acute myocardial infarction. Circulation 2004; 109(25): 3154-7.
113. Orlic D, Kajstura J, Chimenti S, Jakoniuk I, Anderson SM, Li B, et al. Bone marrow cells regenerate infarcted myocardium. Nature 2001; 410(6829): 701-5.
114. Murry CE, Soonpaa MH, Reinecke H, Nakajima H, Nakajima HO, Rubart M, et al. Haematopoietic stem cells do not transdifferentiate into cardiac myocytes in myocardial infarcts. Nature 2004; 428(6983): 664-8.
115. Bel A, Messas E, Agbulut O, Richard P, Samuel JL, Bruneval P, et al. Transplantation of autologous fresh bone marrow into infarcted myocardium: a word of caution. Circulation 2003; 108 (Suppl 1): II247-52.
116. Norol F, Merlet P, Isnard R, Sebillon P, Bonnet N, Cailliot C, et al. Influence of mobilized stem cells on myocardial infarct repair in a nonhuman primate model. Blood 2003; 102(13): 4361-8.
117. Balsam LB, Wagers AJ, Christensen JL, Kofidis T, Weissman IL, Robbins RC. Haematopoietic stem cells adopt mature haematopoietic fates in ischaemic myocardium. Nature 2004; 428(6983): 668-73.
118. Segers VF, Lee RT. Stem-cell therapy for cardiac disease. Nature 2008; 451(7181): 937-42.
119. Bartunek J, Vanderheyden M, Vandekerckhove B, Mansour S, De Bruyne B, De Bondt P, et al. Intracoronary injection of CD133-positive enriched bone marrow progenitor cells promotes cardiac recovery after recent myocardial infarction: feasibility and safety. Circulation 2005; 112(9 Suppl): I178-83.
120. Mansour S, Vanderheyden M, De Bruyne B, Vandekerckhove B, Delrue L, Van Haute I, et al. Intracoronary delivery of hematopoietic bone marrow stem cells and luminal loss of the infarct-related artery in patients with recent myocardial infarction. J Am Coll Cardiol 2006; 47(8): 1727-30.
121. Wall ST, Walker JC, Healy KE, Ratcliffe MB, Guccione JM. Theoretical impact of the injection of material into the myocardium: a finite element model simulation. Circulation 2006; 114(24): 2627-35.
122. Xu X, Xu Z, Xu Y, Cui G. Effects of mesenchymal stem cell transplantation on extracellular matrix after myocardial infarction in rats. Coron Artery Dis 2005; 16(4): 245-55.


{kind=link}
{kind=link}
{kind=link}