










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Links relacionados
Compartilhar

 Permalink
PermalinkRevista Uruguaya de Cardiología
versão On-line ISSN 1688-0420
Rev.Urug.Cardiol. vol.30 no.1 Montevideo abr. 2015
La adventicia arterial: actor olvidado del sistema cardiovascular
Dres. Edmundo I. Cabrera-Fischer1 PhD, Yanina Zócalo2 PhD, Elena C. Lascano1 PhD, Jorge A. Negroni1 PhD, Ing. Sandra Wray1, Dr. Daniel Bia2 PhD
1. Fundación Universitaria Dr. René Favaloro, Buenos Aires, Argentina.
2. Centro Universitario de Investigación, Innovación y Diagnóstico Arterial (CUiiDARTE), Universidad de la República , Uruguay).
Correspondencia: Dr. E. Cabrera-Fischer. Solís 461, Fundación Dr. R. Favaloro, Buenos Aires, Argentina.
Correo electrónico: fischer@favaloro.edu.ar
Financiamiento: Este trabajo se realizó con el subsidio Préstamo BID OC-AR PICT-2008-0340, Buenos Aires, Argentina.
Recibido enero 26, 2015; aceptado marzo 11, 2015.
Resumen
Actualmente se acepta que la adventicia tiene: un importante rol fisiológico al determinar el nivel de nutrición, oxigenación, reparación arterial, regulación de la vasomotricidad, control de la poscarga ventricular, control de la función arterial, etcétera, a la vez que tiene una importante participación en procesos patológicos (por ejemplo, aterosclerosis, hipertensión arterial, génesis de aneurismas de aorta abdominal). Sin embargo, dado lo reciente de la mayoría de los estudios que han redefinido el rol de la adventicia, aún persiste mucho desconocimiento en la comunidad biomédica acerca de la fisiología de la capa adventicia arterial. El presente trabajo tiene como objetivo revisar el rol que actualmente se reconoce para la capa adventicia de la pared arterial.
Palabras clave:
ADVENTICIA
VASA VASORUM
ARTERIAS
Key words:
ADVENTITIA
VASA VASORUM
Introducción
Las teorías empleadas para explicar los principales procesos patológicos vasculares y/o los principales estudios de diagnóstico arterial se centran en las capas más internas de la pared arterial (por ejemplo, medición del espesor íntima-media, función endotelial), dejando de lado la capa adventicia. Igualmente, por décadas en la experimentación de laboratorio las investigaciones han referido casi exclusivamente a la capa íntima y media arterial solamente refiriéndose a la túnica externa para caracterizar aspectos estructurales o relacionados con la función de los vasa vasorum o nervios incluidos en ella. Esta conducta tiene razones bastante antiguas, siendo tal vez la primera de ellas su propio nombre; adventicia proviene del latín, adventicius, que significa “venido de afuera, suplementario”(1,2). Sin embargo, al igual que lo ocurrido con la capa endotelial en los años 80, en los últimos 10 a 15 años ha existido una revalorización de la capa adventicia a partir de estudios experimentales y clínicos.
Actualmente, si bien persiste un gran desconocimiento del rol de la adventicia en la función arterial, se acepta que tiene: 1) un importante rol fisiológico al determinar el nivel de nutrición, oxigenación, reparación arterial, regulación de la vasomotricidad, control de la poscarga ventricular, control de la función arterial, etcétera, a la vez que tiene 2) una importante participación en procesos patológicos (por ejemplo, aterosclerosis, hipertensión arterial, génesis de aneurismas de aorta abdominal)(3).
Bases fisiológicas: conformación y funciones de la adventicia
1.1. Adventicia, injuria y reparación arterial
La pared vascular posee una importante capacidad intrínseca para reparar heridas. En condiciones normales, la pared arterial de animales adultos posee muy bajas, casi indetectables, tasas de proliferación de células endoteliales o de MLV. Sin embargo, si la pared sufre un daño (por ejemplo, por sobreestiramiento) las tasas de proliferación de todas las capas parietales pueden incrementarse incluso más de 100 veces(11-13). Estudios iniciales, principalmente realizados en arterias de roedores, propusieron que las células de MLV de la capa media se encargaban de migrar y diferenciarse en células endoteliales para restablecer la integridad de la capa íntima(11,12). Sin embargo, posteriormente se evidenció en arterias coronarias porcinas, caninas y/o de ratas, que son grupos celulares localizados en la adventicia los que responden inicialmente a la injuria, aumentando su proliferación y migrando fuera de la adventicia, tanto para reparar lesiones de la capa media o de la íntima(14,15). La capacidad de las células adventiciales para migrar a través de la capa media y de la íntima fue evidenciada mediante estudios que han “trasplantado células marcadas” (por ejemplo, con vectores adenovirales) en el lado adventicial de la arteria y les han monitorizado sus movimientos intraparietales, tras generar una injuria arterial en el lado endoluminal(16,17). Entre los tipos celulares que han mostrado proliferar y diferenciarse se encuentran los fibroblasos adventiciales, que pueden convertirse en miofibroblastos, es decir, células con un fenotipo similar al de las células de MLV (por ejemplo, con capacidad sintética de fibras de colágeno)(11,18).
Uno de los eventos tempranos que se requeriría para iniciarse la reparación arterial, inmediatamente luego de la injuria parietal, sería inflamación adventicial con la consecuente acumulación de monocitos y macrófagos activados(19,20). Esta respuesta “local” se requeriría para inhibir a mediadores químicos de la propia adventicia que mantienen “retenido o almacenado” al grupo de células madre/progenitoras, y consecuentemente (tras inhibirlo), posibilitar la “liberación o movilización” de las células progenitoras para alcanzar los destinos necesarios para reparar la pared vascular. La inhibición del reclutamiento leucocitario temprano que forma parte de la respuesta inflamatoria adventicial ha mostrado (tras una injuria) reducir la reparación de la capa íntima(19).
En este contexto, se ha propuesto por Majeski y colaboradores que la adventicia podría tener dos roles diferentes, no mutuamente excluyentes, en la formación de la neoíntima, que se entiende parte de los mecanismos de reparación arterial: (1) actuar como un centro o eje en el proceso de reclutamiento de células inflamatorias que pueden estimular la proliferación y/o migración de células de MLV de la capa media hacia la íntima, así como también reducir las señales que inhiben al nicho de células madre/progenitoras a proliferar y migrar hacia la capa media y/o íntima, para transformarse en otros tipos celulares (por ejemplo, miofibroblastos); y (2) actuar como una fuente (“reserva”) de células precursoras de células neointimales que contribuyen a reparar las lesiones de la capa íntima(11,21).
1.2. Adventicia e irrigación de la pared vascular
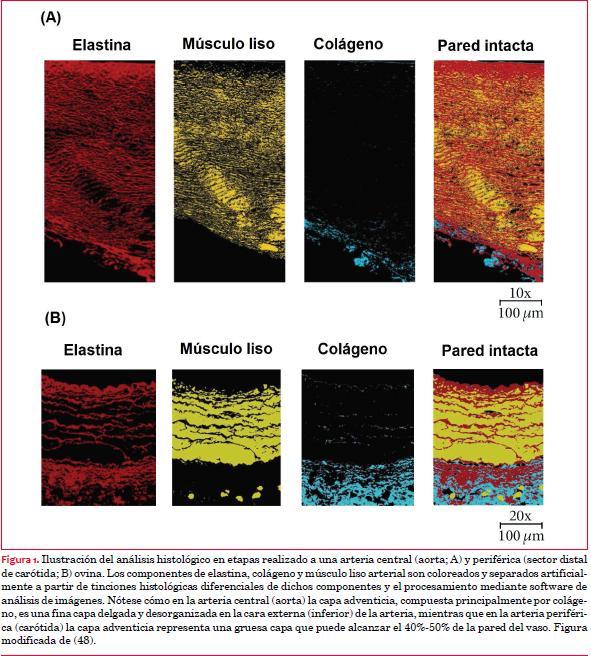
Se podría considerar que los vasa vasorum serían un equivalente de las arterias coronarias en los vasos sanguíneos. En efecto, en ambos casos “coronan” el elemento al cual nutren, a diferencia de lo que ocurre en cualquier otro órgano. La presencia de los vasa vasorum (dativo del latín que traducido significa “vasos de los vasos”) depende del grosor de la pared vascular. Según describiera Wolinsky, las arterias precisan de irrigación parietal propia, provista por los vasa vasorum, a partir de un espesor que contenga más de 29 unidades laminares elásticas en la capa media (aproximadamente 0,5 mm) (8,24). En consecuencia, si las arterias tienen un espesor parietal menor a 0,5 mm, no es necesario contar con los vasa vasorum pues la nutrición se hace por imbibición desde el lumen arterial; sobrepasado dicho grosor parietal conviven las dos alternativas señaladas. Como ejemplo, cuando las arterias intercostales caninas (fuente de la vasa vasorum para la aorta descendente) son ligadas, la túnica media aórtica comienza a sufrir necrosis, evidenciando que mientras las capas celulares internas de la pared arterial se nutren por difusión, las que ocupan posiciones medias y externas intraparietales en la aorta descendente se nutren a partir de la vasa vasorum adventicial(25,26).
Debe recordarse que todo lo descrito sobre la función endotelial no debe ser limitado a la capa de endotelio que está en contacto con la sangre de un vaso de considerable tamaño. Por el contrario, los vasa vasorum de la adventicia y sus prolongaciones a la capa media por su condición vascular poseen una monocapa de endotelio que funciona en forma similar al que se encuentra en la túnica íntima. Por tanto, alteraciones endoteliales pueden afectar a la vasa vasorum adventicial con la consecuente alteración en la capacidad de nutrir y oxigenar a las capas parietales. Al respecto, la disminución de la capacidad de vasodilatación de los vasa vasorum en animales hipertensos contribuiría a la necrosis de la media y disección aórtica(30,31).
1.3. Adventicia y control de la conducta biomecánica arterial: de un rol “pasivo” a un rol “activo y dinámico”
La adventicia contiene colágeno y otros elementos conectivos que posibilitan generar altos niveles de rigidez cuando la presión intraluminal que distiende a la arteria aumenta considerablemente(32,33). Su desempeño ha sido homologado a la cubierta de un neumático, el cual, por más que se le inyecte aire a presión, no va a deformarse sino que aumentará su rigidez. Al respecto, la adventicia, ante elevados niveles de presión sanguínea, hace frente a más del 50% de la carga, por lo que es considerada una capa de “seguridad” contra la ruptura de la pared arterial(34).
Esta visión de que la adventicia desempeña casi exclusivamente un rol mecánico “pasivo” ha cambiando en los últimos años a partir de evidencias que reconocen su rol activo y dinámico en la determinación de la conducta biomecánica de la pared arterial. Al respecto, desde los trabajos experimentales de Soltis y Casis, en 1991, en los que se encontró que la grasa periadventicial atenuaba la respuesta vascular a la norepinefrina en anillos aórticos aislados de rata, el rol regulatorio (activo) de la adventicia ha sido aceptado(35). Al día de hoy, la función paracrina de la adventicia ha sido mostrada en distintas publicaciones y se la relaciona con el control del tono del MLV de la túnica media, el cual estaría bajo el control de las terminaciones nerviosas adventiciales y de mediadores liberados por el tejido adiposo adventicial y periadventicial(36). Al respecto, el ADRF (adventitium-derive relaxing factor), denominado así por analogía con el EDRF (endotelial-derive relaxing factor), ha sido señalado como un regulador clave del tono del MLV(33). La secreción de ADRF ocasiona relajación muscular y vasodilatación arterial independiente de la generada por el óxido nítrico endotelial y la producida por la inervación arterial(37).
A partir de datos experimentales que indicaban la existencia de un factor vasorrelajante adventicial, nuestro grupo emprendió trabajos experimentales destinados a desentrañar la importancia fisiológica que este control activo adventicial tendría sobre la conducta biomecánica arterial. Estos trabajos, realizados en arterias elásticas (tronco braquiocefálico) ovinas mediante estudios in vivo e in vitro (simuladores hemodinámicos cardiovasculares), permitieron demostrar que los niveles de viscoelasticidad parietales de arterias elásticas centrales, bajo condiciones de normopresión, están fuertemente controlados por la capa adventicial de una manera MLV-dependiente. La eliminación de la adventicia determinaba importantes cambios en viscoelasticidad solo en los estudios en los que la reactividad del MLV se encontraba conservada(38); la impedancia y rigidez arterial aumentaron tras eliminar la adventicia únicamente en los experimentos in vivo. Estos resultados permitieron postular que la adventicia, en la condición in vivo y durante valores de presión y deformación basales, participaría en la regulación activa de los niveles de viscoelasticidad y que esto lo llevaría a cabo mediante el control de los valores de respuestas elástica y viscosa y del diámetro arterial. En estas condiciones, la adventicia mantendría reducida la poscarga dinámica ventricular(38). Cabe mencionar que al eliminar la adventicia se evidenció una fuerte constricción muscular, con una significativa reducción del diámetro arterial. Si el papel de la adventicia en las condiciones estudiadas hubiera sido limitar la sobredistensión, la eliminación de la adventicia hubiera conducido a dilatación. Estos resultados, en cuanto a que la eliminación de la adventicia genera constricción, coinciden teóricamente con los trabajos ya mencionados que indican que la adventicia presenta funciones “anticontráctiles”. Finalmente, cabe destacar que los resultados de los estudios in vitro permitieron evidenciar que en estados hemodinámicos basales de normopresión y deformación, el papel pasivo de la adventicia de limitar la sobredistensión arterial no se ejercería. Estos resultados concuerdan con los de Schulze-Bauer y colaboradores(34), quienes encontraron que durante valores de normopresión, la adventicia presenta una elevada distensibilidad y no limita mecánicamente la distensión arterial, mientras que en valores elevados de presión o distensión, la distensibilidad se reduce, limitando en estas condiciones la distensión de la arteria.
Por otra parte, la importancia de una adecuada función/estructura de la capa adventicia, durante la terapéutica con balón de contrapulsación aórtica (CIAO), fue evidenciada en trabajos experimentales adicionales. Al respecto, la realización de CIAO, tanto en condiciones normales como de falla cardíaca aguda, se ha asociado con reducción en la rigidez arterial, es decir, determina una mejoría en los niveles de rigidez arterial, y, consecuentemente, mejora los niveles de poscarga dinámica ventricular y la propia función arterial(7). Por lo tanto, los efectos beneficiosos de la CIAO sobre los niveles de viscoelasticidad arterial no solo dependerían de una adecuada función endotelial, sino también de una adecuada función adventicial(7).
1.4. Adventicia y aterosclerosis
La aterosclerosis, como enfermedad sistémica inflamatoria crónica, es considerada una enfermedad que tras ser generada por una injuria que facilita la activación endotelial, la adhesión monocitaria y la invasión de células inflamatorias, se va propagando (afectando) desde “adentro-hacia-afuera”, es decir desde la capa arterial más interna (capa íntima) progresivamente hacia la capa media y posteriormente externa (adventicia) de la pared arterial. Sin embargo, la dificultad de este modelo para explicar la diferente evolución y vulnerabilidad de las placas de ateroma, sumado a recientes hallazgos que sugieren que la aterosclerosis podría explicarse por un fenómeno “afuera-hacia-adentro”, han llevado a cuestionar el modelo clásico. Bajo esta nueva propuesta, la inflamación podría iniciarse en la adventicia y progresar hacia la capa íntima (39,40). En esta teoría juega un rol clave la vasa vasorum. Numerosos estudios han evidenciado que cambios en la vasa vasorum (por ejemplo, inmadurez de las células de sus paredes, permeabilidad aumentada) o el propio desarrollo descontrolado (angiogénesis intraparietal) estimulado por una noxa (daño parietal) se encuentran relacionados con el desarrollo, progresión y/o la vulnerabilidad de las placas de ateroma. Sin embargo, si la vasa vasorum juega un rol causal o reactivo no está completamente dilucidado. Una revisión detallada de estos aspectos puede encontrarse en la revisión de Kawabe y Hasebe(41) y de Sun(42).
Así como la vasa vasorum adventicial permite llevar oxígeno y nutrientes a los tejidos normales de la pared arterial, también provén de estos requerimientos a los constituyentes celulares de las placas ateroscleróticas en formación. Adicionalmente, la vasa vasorum es la vía de acceso principal a la lesión que utilizan los leucocitos circulantes, y, por lo tanto, cambios en el flujo sanguíneo de la vasa vasorum podrían afectar el crecimiento y la composición de la placa. A manera de ejemplo, diversos inhibidores de la angiogénesis (por ejemplo, endostatina) han mostrado inhibir la progresión de las placas de ateroma, al igual que inhibidores de la proliferación y migración celular.
El remodelado de la pared vascular que se genera ante cambios crónicos en el flujo sanguíneo depende de la interacción de la capa endotelial con las células de las capas media y adventicia(11). Empleando un modelo en que se reduce el flujo carotídeo en ratones, Zhou y colaboradores mostraron que el remodelado (inward) requiere la temprana acumulación por proliferación en la adventicia de macrófagos CXCR3-positivos(20), cuyo rol sería reclutar monocitos hacia la capa adventicial(20). De manera similar, Tang y colaboradores mostraron que la eliminación de macrófagos adventiciales atenúa o elimina el remodelamiento (inward) determinado por reducciones en el flujo sanguíneo carotídeo de ratones(19). Adicionalmente, se ha evidenciado en ratones que paredes arteriales carentes de linfocitos B y T no presentan el remodelado esperable de encontrar cuando se genera hipertensión arterial farmacológica (por ejemplo, tras administrar angiotensina II); remodelado que logra generarse al restablecer en la pared arterial los linfocitos T (no B)(11).
1.6. Adventicia y cirugía vascular/revascularización
Las fístulas arteriovenosas para realizar hemodiálisis frecuentemente presentan problemas que determinan la no viabilidad del cortocircuito arteriovenoso por obstrucciones que dificultan su flujo, principalmente debidas a la generación de hiperplasia intimal. Estos procesos de hiperplasia neointimal estenosante han sido relacionados con la adventicia, la cual estaría involucrada en un proceso inflamatorio concomitante(43). En este proceso, desencadenado por múltiples factores (por ejemplo, biomecánicos hemodinámicas) habría migración de fibroblastos desde la adventicia que estarían en la génesis del proceso de hiperplasia neointimal(44). Ligado a lo anterior, en la cirugía de revascularización coronaria directa se ha asociado la preservación de los vasa vasorum con la viabilidad del injerto(45,46). Souza y colaboradores demostraron un aumento en la viabilidad de los injertos venosos usados para revascularización miocárdica cuando se preserva la adventicia(47).
Conclusión
En el presente trabajo se revisó la literatura científica en aras de brindar información acerca de aspectos fisiológicos y fisiopatológicos de la túnica adventicia arterial. Del análisis de la bibliografía surge que la adventicia funciona como un verdadero centro de reparación, integración, almacenamiento y liberación de reguladores de la función de la pared vascular.
Bibliografía
1. Cabrera Fischer EI. La adventicia: estado actual del conocimiento. Rev Argent Cardiol 2008; 76(1): 58-63.
2. Pagano PJ, Gutterman DD. The adventitia: the outs and ins of vascular disease. Cardiovasc Res 2007; 75(4):636-9.
3. Maiellaro K, Taylor WR. The role of the adventitia in vascular inflammation. Cardiovasc Res 2007; 75(4):640-8.
4. Bia D, Cabrera Fischer EI, Zócalo Y, Armentano RL. The endothelium modulates the arterial wall mechanical response to intra-aortic balloon counterpulsation: in vivo studies. Artif Organs 2011; 35(9):883-92.
5. Cabrera Fischer E, Bia D, Zócalo Y, Camus J, de Forteza E, Armentano R. Effects of removing the adventitia on the mechanical properties of ovine femoral arteries in vivo and in vitro. Circulation Journal 2010; 74(5):1014-22.
6. Cabrera-Fischer E, Bia D, Camus J, Zócalo Y, De Forteza E, Armentano R. Adventitia-dependent mechanical properties of brachio cephalico vine arteries in in vivo and in vitro studies. Acta Physiol (Oxf) 2006; 188(2):103-11.
7. Cabrera-Fischer EI, Bia D, Zócalo Y, Wray S, Armentano R. The adventitia layer modulates the arterial wall elastic response to intra-aortic counterpulsation: in vivo studies. Artif Organs 2013; 37(12):1041-8.
8. Wolinsky H, Glagov S. Comparison of abdominal and thoracic aortic medial structure in mammals. Deviation of manfromthe usual pattern. Circ Res 1969; 25:677-86.
9. Stenmark KR, Nozik-Grayck E, Gerasimovskaya E, Anwar A, Li M, Riddle S, et al. The adventitia: essential role in pulmonary vascular remodeling. Compr Physiol 2011;1(1):141-61.
10. Stenmark KR, Yeager ME, El Kasmi KC, Nozik-Grayck E, Gerasimovskaya EV, Li M, et al. The adventitia: essential regulator of vascular wall structure and function. Annu Rev Physiol 2013;75:23-47.
11. Majesky MW, Dong XR, Hoglund V, Mahoney WM Jr, Daum G. The adventitia: a dynamic interface containing resident progenitor cells. Arterioscler Thromb Vasc Biol 2011;31(7):1530-9.
12. Clowes AW, Reidy MA, Clowes MM. Kinetics of cellular proliferation after arterial injury: I: smooth muscle growth in the absence of endothelium. Lab Invest 1983;49:327-33.
13. Reidy MA, Clowes AW, Schwartz SM. Endothelial regeneration V: inhibition of endothelial regrowth in arteries of rat and rabbit. Lab Invest 1983;49(5):569-75.
14. Holifield B, Helgason T, Jemelka S, Taylor A, Navran S, Allen J, et al. Differentiated vascular myocytes: are they involved in neointimal formation. J Clin Invest 1996;97(3):814-25.
15. Shi Y, O’Brien JE, Fard A, Mannion JD, Wang D, Zalewski A. Adventitial myofibroblasts contribute to neointimal formation in injured porcine coronary arteries. Circulation 1996;94(7):1655-64.
16. Li G, Chen SJ, Oparil S, Chen YF, Thompson JA. Direct in vivo evidence demonstrating neointimal migration of adventitial fibroblasts after balloon injury of rat carotid arteries. Circulation 2000; 101(12):1362-5.
17. Rodriguez-Menocal L, St-Pierre M, Wei Y, Khan S, Mateu S, Calfa M, et al. The origin of post-injury neointimal cells in the rat balloon injury model. Cardiovasc Res 2009;81(1):46-53.
18. Hu Y, Zhang Z, Torsney E, Afzal AR, Davison F, Metzler B, et al. Abundant progenitor cells in the adventitia contribute to atherosclerosis of vein grafts in ApoE-deficient mice. J Clin Invest 2004;113(9): 1258-65.
19. Tang PCY, Qin L, Zielonka J, Zhou J, Matte-Martone C, Bergaya S, et al. MyD88-dependent superoxide-initiated inflammation is necessary for flow-mediated inward remodeling of conduit arteries. J Exp Med 2008; 205(13):3159-71.
20. Zhou J, Tang PC, Qin L, Gayed PM, Li W, Skokos EA, et al. CXCR3-dependent accumulation and activation of perivascular macrophages is necessary for homeostatic arterial remodeling to hemodynamic stresses. J Exp Med 2010;207(9):1951-66.
21. Majesky MW, Dong XR, Hoglund V, Daum G, Mahoney WM Jr. The adventitia: a progenitor cell niche for the vessel wall. Cells Tissues Organs 2012;195(1-2):73-81.
22. Zerwes HG, Risau W. Polarized secretion of a platelet-derived growth factor-like chemotactic factor by endothelial cells in vitro. J Cell Biol 1987; 105(5):2037-41.
23. Alberding JP, Baldwin AL, Barton JK, Wiley E. Effects of pulsation frequency and endothelial integrity on enhanced arterial transmural filtration produced by pulsatile pressure. Am J Physiol Heart Circ Physiol 2005;289(2):H931-7.
24. Wolinsky H, Glagov S. Nature of species differences in the medial distribution of aortic vasa vasorum in mammals. Circ Res 1967; 20(4):409-21.
25. Stefanadis CI, Karayannacos PE, Boudoulas HK, Stratos CG, Vlachopoulos CV, Dontas IA, et al. Medial necrosis and acute alterations in aortic distensibility following removal of the vasa vasorum of canine ascending aorta. Cardiovasc Res 1993; 27(6):951-6.
26. Fuster V, Moreno PR, Fayad ZA, Corti R, Badimon JJ. Atherothrombosis and high-risk plaque: part I, evolving concepts. J Am Coll Cardiol 2005; 46(6):937-54.
27. Ritman EL, Lerman A. The dynamic vasa vasorum. Cardiovasc Res 2007; 75(4):649-58.
28. Moreno PR, Purushothaman KR, Fuster V, Echeverri D, Truszczynska H, Sharma SK, et al.Plaque neovascularization is increased in ruptured atherosclerotic lesions of human aorta: implications for plaque vulnerability. Circulation 2004; 110(14):2032-8.
29. Corti R, Fayad ZA, Fuster V, Worthley SG, Helft G, Chesebro J, et al. Effects of lipid-lowering by simvastatin on human atherosclerotic lesions: a longitudinal study by high-resolution, noninvasive magnetic resonance imaging. Circulation 2001; 104(3):249-52.
30. Hu Y, Xu Q. Adventitial biology: differentiation and function. Arterioscler Thromb Vasc Biol 2011; 31(7): 1523-9.
31. Barsky SH, Rosen S. Aortic infarction following dissecting aortic aneurysm. Circulation 1978; 58(5): 876-81.
32. Liu Y, Dang C, García M, Gregersen H, Kassah GS. Surrounding tissues affect the passive mechanics of the vessel wall: theory and experiment. Am J Physiol 2007; 293(6):3290-300.
33. Sommer G, Regitnig P, Költringer L, Holzapfel GA. Biaxial mechanical properties of intact and layer-dissected human carotid arteries at physiological and supraphysiological loadings. Am J Physiol Heart Circ Physiol 2010; 298(3):H898-912.
35. Soltis EE, Cassis LA. Influence of perivascular adipose tissue on rat aortic smooth muscle responsiveness. Clin Exp Hypertens 1991; 13(2):277-96.
36. Verlohren S, Dubrovska G, Tsang SY, Essin K, Luft FC, Huang Y, et al. Visceral periadventitial adipose tissue regulates arterial tone of mesenteric arteries. Hypertension 2004; 44(3):271-6.
37. Fernandez-Alonso MS. Regulation of Vascular Tone:the fat connection. Hypertension 2004; 44(3):255-6.
38. Bia D, Zócalo Y; Armentano RL, Camus J, De Forteza E, Cabrera-Fischer E. La adventicia reduce la poscarga dinámica ventricular izquierda mediante mecanismos dependientes de la activación muscular lisa. Rev Esp Cardiol 2007; 60(5):501-9.
39. Doyle B, Caplice N. Plaque neovascularization and antiangiogenic therapy for atherosclerosis. J Am Coll Cardiol 2007; 49(21):2073-80.
40. Mulligan-Kehoe MJ. The vasa vasorum in diseased and non-diseased arteries. Am J Physiol Heart Circ Physiol 2010; 298(2):H295-305.
41. Kawabe J, Hasebe N. Role of the vasa vasorum and vascular resident stem cells in atherosclerosis. Biomed Res Int 2014;2014:701571.
42. Sun Z. Atherosclerosis and atheroma plaque rupture: normal anatomy of vasa vasorum and their role associated with atherosclerosis. Scientific World Journal 2014;2014:285058.
43. Diskin CJ. Novel insights into the pathobiology of the vascular access - do they translate into improved care? BloodPurif 2010; 29(2):216-29.
44. Hakim RM, Himmelfarb J. Hemodialysis access failure: a call to action—revisited. Kidney Int 2009; 76(10):1040-8.
45. Tsui JC, Souza DS, Filbey D, Karlsson MG, Dashwood MR. Localization of nitric oxide synthase in saphenous vein grafts harvested with a novel “no-touch” technique: potential role of nitric oxide contribution to improved early graft patency rates. J Vasc Surg 2002; 35(2):356-62.
46. Dashwood MR, Anand R, Loesch A, Souza DS. Hypothesis: a potential role for the vasa vasorum in the maintenance of vein graft patency. Angiology 2004; 55(4):385-95.
47. Souza DS, Johansson B, Bojö L, Karlsson R, Geijer H, Filbey D, et al. Harvesting the saphenous vein with surrounding tissue for CABG provides long-term graft patency comparable to the left internal thoracic artery: results of a randomized longitudinal trial. J Torca Cardiovasc Surg 2006; 132(2): 373-8.


{kind=link}