Saldanha Sylvia1, Boggiano Pablo2, Cadenazzi Mónica2
1Estación Experimental Salto. Facultad de Agronomía, Universidad de la República. Ruta 31 km 21,500 San Antonio. Salto 50009. Uruguay.
Correo electrónico: sylsal71@adinet.com.uy
2Facultad de Agronomía, Estación Experimental Dr. M. A. Cassinoni. Facultad de Agronomía. Universidad de la República.
]]>
Recibido: 23/11/10 Aceptado: 8/1/12
Resumen
]]>
Para comprender mejor las consecuencias de la aplicación de diferentes intensidades de pastoreo, se desarrollaron funciones de respuesta de la producción y composición del forraje de una pastura de Lolium perenne a ofertas de forraje (OF) y se analizaron en forma multivariada las características estructurales. Las OF evaluadas fueron 2,0- 4,5-7,0 y 9,5 kg MS/100 kg de peso vivo animal/día. Las relaciones entre producción de forraje y utilización relativa del forraje con las OF fueron lineales: producción de forraje (kg MS/ha) = 3619+1078OF (P<0,0038; r2=0,74); utilización del forraje (porcentaje) = 88,8-3,86OF (P<0,0066; r2=0,69), no detectándose diferencias en el forraje desaparecido. La relación de la OF de la biomasa aérea pre pastoreo y la altura del follaje para todo el período fue: biomasa aérea (kg MS/ha) = -758+237x, (P<0,0001; r2=0,79) siendo x = altura (cm). La biomasa de láminas prepastoreo fue afectada linealmente (P<0,01; r2=0,70) por la biomasa de láminas verdes remanentes del pastoreo anterior y por la densidad de macollos (P<0,0001; r2=0,67). El análisis de componentes principales indicó que bajo esas condiciones, únicamente la mayor OF evaluada mantenía los valores de las variables relativamente altos, sugiriendo así una mayor productividad y estabilidad de la pastura bajo este manejo. Las relaciones obtenidas se consideran útiles para el manejo de las pasturas, a pesar de que las condiciones ambientales variaron durante el período de medición, la frecuencia del pastoreo fue mayor a la recomendada y algunos macollos pasaron al estado reproductivo.
Palabras clave: RAIGRÁS PERENNE, INTENSIDAD DEL PASTOREO, BIOMASA DE LÁMINAS
]]>
Summary
Herbage Allowance, Production and Composition of a Lolium Perenne Pasture
For a better understanding of the consequences of the application of different grazing intensities, functions of response of composition and forage production of a pasture of Lolium perenne under different grazing herbage allowance levels (OF) were developed, and their structural characteristics were analysed with a multivariate method. The OF evaluated were 2.0- 4.5-7.0 and 9.5 DM kg/100 kg of animal liveweight/day. The relationships between forage production and relative forage utilization with the OF were lineal: forage production (DM kg/ha)=3619+1078OF, (P<0.0038; r2=0.74); forage utilization (percentage )=88.8-3.86OF, (P<0.0066; r2=0.69), but no differences were detected in the disappeared forage. The relationship between pre-graze aerial biomass and foliage height (cm) for the entire period was: aerial biomass (DM kg/ha) = -758 + 237x (P<0.0001; r2 = 0.79), for x=height (cm). The pre-graze green leaves biomass was lineally affected (P<0.01; r2=0.70) ) by the remainder green leaves biomass post-graze and by tillers density (P<0.0001; r2=0.67). Principal component analysis indicated that, under these conditions, only the biggest OF evaluated maintained the values of variables relatively high, suggesting a highest pasture productivity and stability under these treatments. The relationships achieved are useful for pasture management, although the environmental conditions changed during the experimental period, the grazing frequency was higher than the recommended and some tillers began to reproductive state.
Key words: PERENNIAL RYEGRASS, GRAZING INTENSITY, LEAVES BIOMASS
]]>
Introducción
El manejo de las pasturas es complejo. El ajuste de la presión de pastoreo es una herramienta importante para obtener una eficiente utilización (Leaver y Weissbach, 1993); sin embargo en general la intensidad del pastoreo en Uruguay es excesiva afectando la productividad y persistencia de las pasturas.
Hay especies que son capaces de adaptarse al ambiente y al manejo de la defoliación a través de modificaciones progresivas y reversibles de las características estructurales de forma de restablecer el equilibrio entre demanda y suministro de recursos (Chapman y Lemaire, 1993; Lemaire y Chapman, 1996). Lolium perenne es una especie de gran plasticidad fenotípica, ya que presenta similares producciones de forraje bajo diferentes intensidades de pastoreo continuo y condiciones ambientales no limitantes (Matthew et al., 2000; Gastal et al., 2004; Agnusdei y Assuero, 2004). Pero estos procesos difieren bajo pastoreos intermitentes y condiciones ambientales limitantes y/o variables, y las modificaciones en la estructura no compensan los efectos ocasionados por el manejo (Agnusdei y Assuero, 2004; Saldanha et al., 2010).
]]>La estructura de la pastura determina su productividad primaria y secundaria al afectar su crecimiento y posibilidad de utilización por el animal. Al definir estrategias de manejo, se debe considerar además de la producción de forraje otras características que afectan la accesibilidad del forraje por parte del animal: la biomasa aérea, la altura del follaje (variable que determina el tamaño de bocado del animal), la biomasa verde y el tamaño de las láminas (Prache y Peyraud, 1997). El conocer las relaciones entre variables estructurales, como por ejemplo biomasa aérea y altura, facilita la implementación práctica de las decisiones de manejo.
El objetivo de este trabajo fue mejorar la comprensión y predicción de las consecuencias de la aplicación de diferentes intensidades de pastoreo, desarrollando funciones de respuesta de la producción y composición del forraje de una pastura de Lolium perenne a la intensidad de pastoreo, aplicadas como ofertas de forraje (OF) durante la primavera, y evaluar la estabilidad de la pastura de continuar aplicándose los pastoreos, a través de un análisis de componentes principales de todas las variables estudiadas.
Materiales y métodos
]]>
El área experimental se localizó en la Estación Experimental Mario A. Cassinoni de la Facultad de Agronomía de la Universidad de la República Oriental del Uruguay (Latitud, 32º20’S; Longitud 58º20’W, a 61 m sobre el nivel del mar), Paysandú. La pastura utilizada fue una mezcla de Lolium perenne cv Horizon, Trifolium repens cv Zapicán y Lotus corniculatus cv San Gabriel sembrada en directa el 30 de abril de 2006 con 15, 2 y 8 kg de semilla/ha respectivamente, sobre un Brunosol eútrico típico háplico de la Unidad San Manuel. La gramínea se sembró en línea a 17 cm y se fertilizó a la siembra con 100 kg/ha de 18-46-0 y el 30 de mayo con 50 kg/ha de 46-0-0. La presencia de leguminosas y malezas durante el período abarcado por este estudio, fue ínfima. Los datos meteorológicos (precipitaciones, temperaturas medias, máxima medias y mínimas medias y evaporación del tanque A) se obtuvieron del Aeropuerto Chakling. Se elaboró un balance hídrico decádico del suelo y se calculó la temperatura media acumulada desde la siembra al primer pastoreo y entre pastoreos. Las condiciones ambientales fueron descriptas por Saldanha et al. (2010).
Los tratamientos consistieron en cuatro OF: 2,0; 4,5; 7,0 y 9,5 kg de MS/100 kg del peso vivo animal que se aplicaron el 6 de agosto, 20 de septiembre, 22 de octubre, 10 de noviembre y 30 de noviembre, ya que el área experimental fue parte de un proyecto en el que los animales pastoreaban franjas diarias (cuya área se estimaba en función de la biomasa aérea presente) y retornaban a ella en cada ciclo de pastoreo. Por esto la frecuencia de pastoreo si bien fue la misma para todos los tratamientos difirió en el período de estudio (45, 32,19 y 20 días de descanso). Se utilizaron seis novillos tester fijos en cada OF, más animales volantes para ajustar la misma. El diseño experimental fue completamente al azar con dos repeticiones en cada área, utilizándose ocho parcelas de diferente superficie según tratamientos, para mantener similar densidad animal (Shaw et al., 1985; Giesbrecht, 1989).
Para el análisis estadístico de los datos fueron utilizadas técnicas de regresión. Se ajustaron modelos de regresión lineales y cuadráticos para cada fecha entre las OF y las variables estudiadas, comparándose las funciones resultantes. En el caso en que las funciones no eran estadísticamente diferentes entre fechas, se ajustó una función general para todo el período. En todos los casos se estudió el comportamiento de los errores para los modelos ajustados, mediante técnicas de diagnóstico gráficas y numéricas. El criterio de significación utilizado en los ajustes de modelos de regresión fue de 6%.
El modelo usado fue:
Yijkl= m +Oj + ξil + MMk + (MM x O)jk + εijkl
Donde:
Yijkl es la variable;
m es la media poblacional;
]]> Oj es el efecto de la OF;ζil es el error de la OF;
MMk es el efecto del período transcurrido entre las mediciones; (MM x O)jk es la interacción entre el momento de medición y la OF;
εijkl es el error experimental en el cual se modeló la autocorrelación existente entre las unidades medidas en diferentes momentos.
Se realizó un análisis de componentes principales con todas las variables determinadas en este trabajo y en el de Saldanha et al. (2010). Se estudió la evolución en el tiempo de los valores de las variables en forma multivariada con el objetivo de evaluar la estabilidad de los resultados de continuar aplicándose los pastoreos.
Determinaciones
Previo a cada pastoreo se extraían tres muestras del forraje disponible por parcela, previa medición de la altura del follaje en su posición natural (3 a 5 puntos/muestra) (Rhodes y Collins, 1993). Las muestras consistían en un pan de tierra de 20 cm x 17 cm y de 10 cm de profundidad. Se les extraía la tierra y se lavaban las raíces (Mannetje, 2000), con la finalidad de poder contabilizar el número de plantas (variable analizada en otro trabajo).
Después de cada pastoreo se medía con regla la altura del forraje rechazado en cinco muestras por parcela de 20 x 17 cm de superficie, luego de lo cual se cosechaba el forraje de cada muestra con tijera de aro al ras del suelo.
Variables
]]>En cada muestra pre pastoreo, luego de eliminar las raíces, contar el número de plantas y macollos (variables no analizadas en este trabajo) y clasificarlos subjetivamente (en función de su diámetro) en dos categorías: grandes y pequeños, se procedía a separar con tijera el material aéreo senescente (seco) del material verde. A este último se lo fraccionaba en sus componentes: láminas expandidas, láminas en expansión y pseudotallos (total de biomasa de vainas) (Grant, 1993). Se contabilizaba cada fracción y se colocaba en estufa hasta peso constante. Así se determinó la biomasa aérea prepastoreo como la suma de la biomasa muerta y la biomasa verde (suma de todos los componentes verdes: biomasa de láminas expandidas, biomasa de láminas en expansión y biomasa de vainas) (Jewiss, 1993). También se midió la altura del pseudotallo con una regla en 20 macollos grandes y 20 macollos chicos por muestra.
En las muestras del forraje remanente (pos pastoreo) se determinaba su masa, se separaba sus componentes: láminas verdes, vainas y material muerto, y se secaban en estufa hasta peso constante.
La producción de forraje se estimó por diferencia de la biomasa aérea pre pastoreo y la biomasa remanente del pastoreo anterior (Frame, 1993; Shaw et al., 1985). La utilización relativa por los animales de la biomasa presente en cada pastoreo se calculó a través de la diferencia de biomasa pre y pos pastoreo (forraje desaparecido) dividida la biomasa pre pastoreo (Shaw et al., 1985). Estas variables se analizaron para todo el período en estudio y no por fecha.
]]>
Resultados y discusión
Producción de forraje, forraje desaparecido y utilización relativa del forraje
La producción de forraje total desde el 7 de agosto al 30 de noviembre ajustó a un modelo lineal (P<0,0038; r2=0,74). La función fue: producción de forraje (kgMS/ha) = 3619 + 1078,5 OF. Sin embargo no existió ajuste, ni se detectaron diferencias, en el forraje desaparecido (6228 ± 1701 kg MS/ha) entre las OF. Esto último se debió a que la utilización relativa del forraje disminuyó al aumentar las OF (P<0,0066), siendo la ecuación de predicción obtenida: utilización del forraje (%) = 88,8 - 3,86 OF, con un r2 = 0,69.
La altura del forraje remanente fue en promedio del período de 3,0 ± 1,5; 6,7 ± 1,7; 10,8 ± 2,0; 11,6 ± 2,7 cm para las OF de 2,0; 4,5; 7,0 y 9,5 % respectivamente. Estos resultados coinciden con el trabajo de Hernández Garay et al. (1999), que evaluaron cinco alturas de cortes (de 2 a 16 cm) encontrando aumentos significativos en la biomasa acumulada hasta 12 cm. También Pérez et al. (2002) alcanzaron la mayor biomasa acumulada al cosechar entre 9 y 12 cm. Las OF evaluadas provocaron diferencias (P<0,0005) en la biomasa de hojas remanentes verdes pos pastoreo (Cuadro 1), reflejando áreas foliares remanentes mayores al crecer la OF. Al quedar con mayor área foliar remanente bajo pastoreos menos intensos, las plantas pueden captar más luz y por lo tanto producir mayor cantidad de fotosintatos, que originan macollos más grandes y mayor producción de biomasa (Skinner y Nelson, 1994).

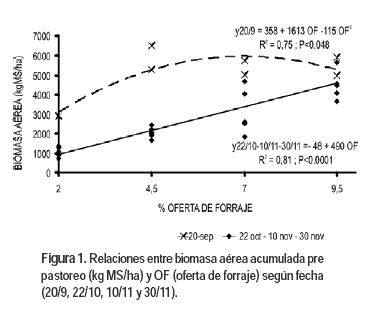
Biomasa aérea
En la Figura 1 se presentan las relaciones entre la biomasa aérea y las OF. Existió interacción entre OF y el momento de la medición ya que se observa que el 20 de setiembre la función ajustó a un modelo cuadrático con máximo en 7% de OF y para las siguientes fechas la relación fue lineal. Esto indica que al 20 de setiembre, por el amplio período de rebrote (45 días) o por las menores intensidades de luz en invierno con respecto a primavera, se habría alcanzado el IAF (Índice de Área Foliar) óptimo con la OF de 7%. Similar resultado obtuvieron Hernández Garay et al. (2000) presentando una menor fotosíntesis neta por área de suelo y por área foliar la defoliación a 16 cm frente a la de 12 cm. Las OF menores a 7% parten de un área foliar remanente menor, por lo que requieren de más tiempo para alcanzar la tasa máxima de fotosíntesis neta (Parsons y Chapman, 2000) ya que es menor la intercepción de luz (Gastal et al., 2004).
]]>
La relación lineal entre la biomasa aérea pre pastoreo y las OF de los tres últimos pastoreos se explica porque al disminuir el período entre pastoreos la acumulación de biomasa fue menor (P<0,0001) en este período (2767 ± 1555 kg MS/ha) con respecto al 20 de setiembre (4918 ± 1327 kg MS/ha). Además la pastura en la primavera requiere mayor área foliar para interceptar toda la radiación incidente, ya que aumenta la intensidad de luz y el largo del día (Parsons y Chapman, 2000). A esto se le sumaron cambios en la morfología de las pasturas, por el proceso de alargamiento de entrenudos, que modifican la distribución vertical de las hojas reduciendo el sombreado de las hojas inferiores (Parsons, 1988), manifestándose más el pasaje al estado reproductivo a menor intensidad del pastoreo (Hernández Garay et al., 1997; Parsons y Chapman, 2000). Por lo que, aún en las mayores OF evaluadas, no se logró interceptar el 95% de la radiación incidente (Índice de Área Foliar óptimo).
Altura del forraje
La relación entre la altura del forraje pre pastoreo y las OF al inicio ajustó (r2=0,62) una función cuadrática (P<0,007), siendo la ecuación conjunta para el 20 de setiembre y el 22 de octubre la siguiente: altura del forraje (cm) = -3,7 + 8,4 OF – 0,6 OF2., con máximo en 7% de OF. A medida que aumenta la OF, aumenta el peso de los macollos por un incremento en el peso de láminas y vainas (Saldanha et al., 2010). Pero las láminas más pesadas son más largas y «caen» por su peso curvándose, determinando una reducción en la altura del follaje sin alterar (9,5% de OF). En los pastoreos de noviembre, cuando se dio el máximo desarrollo reproductivo, y el alargamiento de los entrenudos del tallo, la relación (P<0,0003) entre la altura y las OF fue lineal (altura del forraje en cm = 7,3 + 1,4 OF), con un r2=0,63.
Composición de la biomasa
Biomasa verde
La biomasa verde se asocia más con la productividad animal que la biomasa aérea total. La biomasa verde el 20 de setiembre ajustó una función cuadrática (P<0,069, r2=0,70): biomasa verde 20/9 (kg MS/ha)= 504 + 1418 OF – 102 OF2 con un máximo en 6,6% de OF. En los siguientes pastoreos la relación entre la biomasa verde pre pastoreo (P<0,001, r2 = 0,84 y P<0,001, r2 = 0,85 respectivamente) y las OF fue lineal (biomasa verde 22/10 (kg MS/ha) = 692 + 230 OF y biomasa verde 10/11 (kg MS/ha) = - 381 + 418 OF). Al final del período experimental (30 de noviembre) la relación (P<0,075, r2 = 0,87) pasó a ser nuevamente cuadrática: biomasa verde 30/11 (kg MS/ha) = 1419 + 411 OF – 66 OF2, con un mínimo en 3% de OF.
]]>Como se expresó, el 20 de setiembre seguramente se alcanzó con la mayoría de las OF evaluadas el IAF que maximiza la intercepción de la luz incidente, debido a un mayor período de rebrote, y quizás también por menores gastos energéticos de respiración y de fotorespiración dadas las menores temperaturas (Donaghy y Fulkerson, 1998; Lee et al., 2008).
Sin embargo con el transcurso de la primavera el ambiente lumínico cambió. Además la frecuencia de pastoreo aumentó determinando menor crecimiento foliar (Parsons, 1988), el que pudo verse afectado también por la escasez de precipitaciones que limitó el agua disponible en el suelo hasta el 22 de octubre (Saldanha et al., 2010), esto determinó que la biomasa verde fuera incrementando al aumentar la OF pero sin alcanzar un máximo.
Composición de la biomasa verde
]]>
Biomasa de láminas
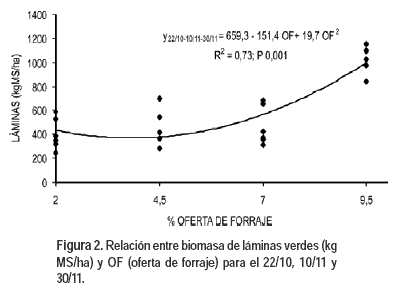
Las láminas de las hojas en general son de mayor calidad y más apetecidas por los animales que las vainas. La biomasa de láminas verdes (Figura 2) presentó un ajuste cuadrático con las OF a partir del 22 de octubre. La ecuación conjunta para las tres fechas fue: biomasa de láminas (kg MS/ha) = 659,3 – 151,4 OF + 19,7 OF2 (P<0,001), siendo explicada el 73% de la variación en la biomasa de láminas por las OF. La mínima acumulación de biomasa de láminas se dio con una OF de 3,8%. Pérez et al. (2002) evaluando esta especie bajo cortes frecuentes a cinco alturas (3 a 15 cm) obtuvieron también que el peso de las láminas disminuye al aumentar la intensidad del corte. Las menores OF presentan menor crecimiento de hojas como resultado de una remoción casi total de las hojas y de un intervalo de pastoreo muy frecuente.
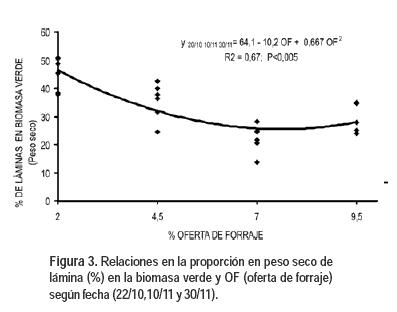
La relación de la proporción en peso seco de láminas en la biomasa verde con las OF (Figura 3) fue también cuadrática desde el 22 de octubre. La ecuación conjunta para las tres fechas fue significativa (P<0,005), con r2=0,66, porcentaje de biomasa de láminas = 64,1 – 10,2 OF + 0,67 OF2, y el mínimo se dio en 7,6% de OF. A menor OF mayor proporción de láminas (menor de vainas) en la biomasa verde. Las menores OF afectaron más el crecimiento de las vainas que el de las láminas, al ser menor el forraje acumulado pre pastoreo y su altura. El aumento en la longitud de las hojas durante el rebrote inicialmente es mayor en la lámina que en la vaina (Matthew et al., 2000). En situaciones de sombreado puede ocurrir alargamiento de entrenudos, de forma de ubicar a las nuevas láminas en estratos superiores (Lemaire, 2001). En cantidades relativas, el pastoreo más intenso proporcionó una mejor composición del forraje expresada como proporción de láminas (P<0,0001), pero en cantidades absolutas la OF de 9,5% ofreció mayor biomasa de láminas que los restantes tratamientos (P<0,0001). Hernández Garay et al. (1999) obtuvieron la mayor proporción de láminas en la biomasa total en los cortes a 2 y 4 cm, no siendo significativamente diferentes entre sí las proporciones de láminas a 8, 12 y 16 cm de altura de corte.
]]> 
Con excepción al 22 de octubre (que coincide con baja disponibilidad de agua en el suelo), en los demás períodos evaluados existió una relación lineal significativa (P<0,0001) y con r2=0,70, entre la biomasa aérea acumulada y la biomasa de láminas, Biomasa aérea (kg MS/ha) = 1482,2 + 1,95 x, siendo x = biomasa de láminas.
La tasa de expansión foliar es afectada por la cantidad de hojas remanentes pos defoliación (Grant et al., 1981). La relación entre la biomasa de lámina pre pastoreo y la biomasa de lámina verde remanente del pastoreo anterior fue significativa (P<0,010; r2=0,70) y lineal: biomasa de lámina pre pastoreo (kg MS/ha) = 302,04 + 1,04 x, siendo x = biomasa de lámina remanente del pastoreo anterior.
La biomasa de láminas depende del número de hojas/macollo, de la densidad de macollos y del tamaño de las láminas.
Número de hojas/macollo
No se encontró relación entre el número de hojas/macollo y las OF aplicadas. El promedio de hojas por macollo de todos los tratamientos y período fue de 2,85 ± 0,25 (Cuadro 2). El número de hojas vivas por macollo para un genotipo, bajo un amplio rango de condiciones estables (ambientales y de manejo), es relativamente constante a través de la vida de la planta según Skinner y Nelson (1994), Chapman y Lemaire (1993), Parsons y Chapman (2000) y Agnusdei y Assuero (2004). Es la resultante del balance entre el número de hojas acumuladas que aparecen y desaparecen (Agnusdei y Assuero, 2004), estando la tasa de aparición de hojas y su longevidad relacionadas (Lemaire y Chapman, 1996; Nelson, 2000; Parsons y Chapman, 2000). Lolium perenne tiene en promedio 3,5 hojas/macollo; cada vez que una nueva hoja se expande la más vieja muere (Mathew et al., 2000, Parsons y Chapman, 2000).
]]> 
Existió variación en el tiempo, variando el número de hojas/macollo entre fechas de pastoreo (Cuadro 2). Luego del primer pastoreo el número de hojas vivas/macollo disminuyó (P<0,0006) tendiendo a aumentar hacia el final del período experimental. El número de hojas/macollo es constante siempre que el intervalo entre defoliaciones sea mayor a la vida media de las hojas y la disponibilidad de nutrientes y agua no sea limitante (Lemaire y Agnusdei, 2000). Para Fulkerson y Slack (1994, 1995) la frecuencia óptima de pastoreo es aquella que permite expandir el número de hojas correspondiente a la vida media de una hoja, es decir antes de que comience la senescencia de la primera hoja producida luego del pastoreo. O sea que es posible medir la frecuencia de los pastoreos en días o considerando el estado fisiológico de las plantas (Lee et al., 2007).
Según Fulkerson y Slack (1994, 1995) Lolium perenne se debería pastorear cuando se desarrollan tres láminas totalmente expandidas/macollo pos defoliación ya que la senescencia comienza en el estado de 3,5 hojas/macollo (Fulkerson y Slack, 1994). Para el presente ensayo, la frecuencia de los pastoreos fue mayor a la óptima, ya que el número de hojas expandidas/macollo promedio del período y de todos los tratamientos fue de 1,74 ± 0,25. La frecuencia entre tratamientos fue similar (tanto medida en días o a través del número de hojas promedio totalmente expandidas por macollo (P<0,12), pero varió la frecuencia de los pastoreos entre períodos, tanto en días como en número de hojas expandidas/macollo (P<0,0007). El pasaje al estado reproductivo que lleva a un menor número de meristemas foliares, también pudo haber influido en la variación del número de hojas/macollo en el tiempo.
Densidad de macollos
La variación en la biomasa de láminas con las OF fue explicada en parte por la variación en la densidad de macollos, la que tuvo un incremento lineal (P<0,001; r2=0,76) de 209 macollos/m2 por cada 1% de incremento en la OF (Saldanha et al., 2010). La relación entre la biomasa de láminas y la densidad de macollos fue lineal en los tres últimos pastoreos, y para el conjunto de éstos fue significativa (P<0,0001) con un r2=0,67, biomasa de láminas (kg MS/ha) = -241,7 + 0,28 x, siendo x = número de macollos/m2.
Peso de lámina
El 22 de octubre (Figura 4) el peso de lámina y la OF ajustó mejor un modelo cuadrático: peso de lámina (mg) = 7,26 – 0,45 OF - 0,05 OF2; (P< 0,044; r2=0,81) con un mínimo en 4,5 % OF.
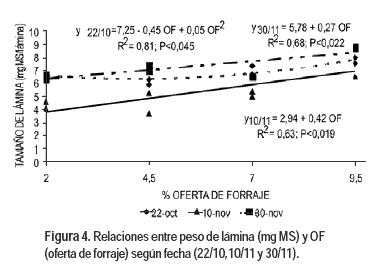
La defoliación afecta el crecimiento foliar (Richards, 1993) posiblemente por la modificación de la altura del pseudotallo (Duru y Ducrocq, 2000). El tamaño de la lámina depende de la tasa de elongación foliar y de la duración de la tasa de expansión de las hojas. Mayores OF implican mayor longitud de vainas remanentes y por lo tanto hojas más largas, debido principalmente a un aumento en la duración del proceso de expansión, más que a un aumento en la tasa de extensión (Duru y Ducrocq, 2000; Gastal et al., 2004). Esto se debe a que la longitud del pseudotallo afecta la calidad de la luz que es percibida por la base de las hojas en crecimiento, y al aumentar el rojo lejano aumenta la longitud de las nuevas hojas (Gastal et al., 2004). Además la tasa de expansión foliar disminuye cuando se remueve parte de las vainas (Grant et al., 1981) por la eliminación de los meristemas intercalares de hoja (Gastal et al., 2004).
]]>
La OF afectó la longitud del pseudotallo (Figura 5). El 20 de setiembre ajustó un modelo cuadrático. En las mayores OF ocurrió alargamiento de entrenudos como forma de mejorar el acceso a la luz de las nuevas láminas (Lemaire, 2001). El 22 de octubre no existió ajuste entre ambas variables, momento que coincidió con el mínimo de agua almacenada en el suelo (Saldanha et al., 2010). En noviembre el ajuste fue lineal, difiriendo las funciones según fecha de pastoreo. Seguramente el pasaje diferencial al estado reproductivo de los macollos, según tamaño y OF incidió en estos resultados.
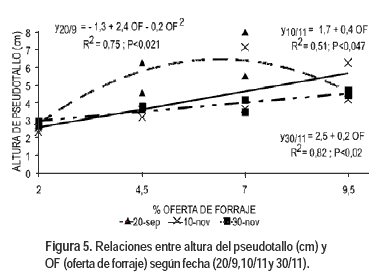
Por esto el ajuste del peso de la lámina (cuya mayor variación está dada por la longitud de la misma) con la longitud del pseudotallo no fue aceptable. Existió una relación lineal significativa (P<0,0001) con un r2=0,70, entre el peso promedio de la lámina ya expandida de los macollos pequeños y la longitud del pseudotallo de este grupo de macollos. Para el conjunto de las todas las fechas fue: peso promedio de lámina ya expandida en macollos pequeños (mg) = - 0,07 + 1,73 x, para x = longitud del pseudotallo de macollos pequeños (cm). La inexistencia de relación entre las variables en los macollos grandes podría estar parcialmente explicada por el alargamiento de los entrenudos, debido al pasaje al estado reproductivo (Skinner y Nelson, 1995). También es necesario considerar que los pesos por unidad de longitud de hoja y de área difieren con la intensidad del corte, con el sombreado y con el estado de desarrollo de la hoja (en expansión o al inicio de la senescencia) (Grant et al., 1981).
No existió relación entre el peso de las láminas y los grados días acumulados entre pastoreos (r2=0,44). En general al aumentar la temperatura hasta 20-25 ºC, las hojas se expanden a mayor velocidad durante un período más corto, siendo su tamaño final mayor (Parsons y Chapman, 2000). Algunos factores que pudieron afectar esta relación, a través de un efecto negativo en las tasas de crecimiento, son la deficiencia de N (Skinner y Nelson, 1995) y/o el déficit de agua en el suelo (Duru y Ducrocq, 2000). Si bien la tasa de expansión foliar es mayor durante el pasaje al estado reproductivo (Lemaire y Chapman, 1996) al alargarse los entrenudos el largo del pseudotallo disminuye afectando más la longitud de la lámina que la de la vaina (Skinner y Nelson, 1995).
]]>Análisis de componentes principales
El estudio multivariado de los valores de las variables estudiadas nos permite visualizar la evolución en el tiempo de los resultados en forma conjunta y no aisladamente cada variable.
Como se observa en la Figura 6, el 76,5% de la variabilidad total queda explicada por los ejes 1 y 2, y el coeficiente de correlación coefenética es alto. Al inicio de la evaluación, las variables en todos los tratamientos presentaron valores altos a excepción de la menor OF. A medida que se suceden los pastoreos, en las OF de 2,0 y 4,5% los valores disminuyen rápidamente y en forma paulatina en la OF de 7,0%. Esta última OF, al final del período de evaluación, presenta valores de las variables, cercanos a los pastoreos más intensos. En estas condiciones, únicamente la mayor OF evaluada mantuvo los valores relativamente altos de las variables, sugiriendo una mayor productividad y estabilidad de la pastura bajo este manejo.
Conclusiones
]]>Las relaciones entre producción de forraje y utilización relativa del forraje con las OF fueron lineales, no detectándose diferencias en el forraje desaparecido. La biomasa de láminas verdes remanentes a los pastoreos fue afectada por las OF, lo que seguramente determinó diferencias en la proporción de luz interceptada y en el forraje acumulado entre pastoreos.
Las relaciones con la OF de la biomasa aérea pre pastoreo y de la altura del follaje variaron con el tiempo, en parte debido a períodos de rebrote diferentes entre pastoreos. Sin embargo ambas variables, biomasa y altura pre pastoreo, ajustaron una función lineal entre sí para todo el período.
La biomasa de láminas prepastoreo fue afectada linealmente por la biomasa de láminas verdes remanentes del pastoreo anterior. La biomasa de láminas se incrementó al aumentar la OF, debido a la variación en la densidad de macollos y en el peso de cada lámina, ya que el número de hojas/macollo no fue afectada por las OF. La proporción de lámina en la biomasa verde ajustó una función cuadrática con un mínimo en 7,6% de OF, lo que pudo estar incidiendo en la proporción del forraje utilizado por los animales
El análisis de componentes principales indicó que bajo esas condiciones únicamente la mayor OF evaluada mantiene los valores relativamente altos de las variables, sugiriendo una mayor productividad y estabilidad de la pastura bajo este manejo.
Bibliografía
Agnusdei MG, Assuero SG. 2004. Leaf tissue flows under grazing and sward structure of different temperate forage grasses in the humid pampas of Argentina [CD-ROM]. En: II Symposium in Grassland Ecophysiology and Grazing Ecology; Curitiba, Paraná, Brasil. Curitiba: Universidad de Curitiba.
Chapman DF, Lemaire G. 1993. Morphogenetic and structural determinants of plant regrowth after defoliation [CD-ROM]. En: Proceedings of the 17th International Grassland Congress. Wellington : SIR Publishing. pp. 55 - 64.
]]>Donaghy DJ, Fulkerson WJ. 1998. Priority for allocation of water-soluble carbohydrate reserves during regrowth of Lolium perenne. Grass and Forage Science, 53:211 - 218.
Duru M, Ducrocq H. 2000. Growth and senescence of the successive grass leaves on a Cocksfoot tiller. Effect of nitrogen and cutting regime. Annals of Botany, 85: 645 - 653.
Frame J. 1993. Herbage mass. En: Davies A, Baker RD, Grant SA, Laidlaw AS. [Eds.]. Sward Measurement Handbook. Reading: The British Grassland Society. pp 39 - 67.
Fulkerson WJ, Slack K. 1995. Leaf number as a criterion for determining defoliation time for Lolium perenne: 2. Effect of defoliation frecuency and height. Grass and Forage Science, 50: 16 - 20.
Fulkerson WJ, Slack K. 1994. Leaf number as a criterion for determining defoliation time for Lolium perenne: 1. Effect of water-soluble carbohydrates and senescence. Grass and Forage Science, 49: 373 - 377.
]]>Gastal F, Lemaire G, Lestienne F. 2004. Defoliation, shoot plasticity, sward structure and herbage utilisation [CD-ROM]. En: II Symposium in Grassland Ecophysiology and Grazing Ecology; Curitiba, Paraná, Brasil. Curitiba : Universidad de Curitiba.
Giesbrecht FG. 1989. Experimental design and statistical inference : Generalized least squares and repeated measures over time. En: Marten GC [Ed.]. Grazing research : design, methodology and analysis. Madison : Crop Science Society of America. pp. 85 - 96.
Grant S. 1993. Resource description : vegetation and sward components. En: Davies A, Baker RD, Grant SA, Laidlaw AS. [Eds.]. Sward Measurement Handbook. Reading: The British Grassland Society. pp. 69 - 97.
Grant SA, Barthram GT, Torvell L. 1981. Components of regrowth in grazed and cut Lolium perenne swards. Grass and Forage Science, 36: 155 - 168.
Hernández Garay A, Hodgson J, Matthew C. 2000. The influence of defoliation height on dry matter partitioning and CO2 exchange of perennial ryegrass miniature swards. Grass and Forage Science, 55: 372 - 376.
]]>Hernández Garay A, Hodgson J, Matthew C. 1999. Tiller size/density compensation in perennial ryegrass miniature swards subject to differing defoliation heights and a proposed productivity index. Grass and Forage Science, 54: 347 - 356.
Hernández Garay A, Hodgson J, Matthew C. 1997. Effect of spring grazing management on perennial ryegrass and ryegrass-white clover pastures : 2. Tiller and growing point densities and population dynamics. New Zealand Journal of Agricultural Research, 40: 37 - 50.
Jewiss OR. 1993. Shoot development and number. En: Davies A, Baker RD, Grant SA, Laidlaw AS. [Eds.]. Sward Measurement Handbook. Reading : The British Grassland Society. pp. 99 - 120.
Leaver JD, Weissbach F. 1993. Trends in intensive temperate grassland systems. En: Proceedings of the 17th International Grassland Congress. Wellington : SIR Publishing. pp.1481 - 1485.
Lee JM, Donaghy DJ, Roche JR. 2008. Effect of defoliation severity on regrowth and nutritive value of perennial ryegrass dominant swards. Agronomy Journal, 100: 308 - 314.
]]>Lee JM, Donaghy DJ, Roche JR. 2007. Effect of grazing severity and fertiliser application during winter on herbage regrowth and quality of perennial ryegrass (Lolium perenne L.). Australian Journal of Experimental Agriculture, 47: 825 - 832.
Lemaire G. 2001. Ecophysiology of grassland : Dynamic aspects of forage plant populations in grazed swards [CD-ROM]. En: Proceedings of the XIX International Grassland Congress; 11 - 21 febrero 2001; Sao Paulo, Brasil.
Lemaire G, Agnusdei M. 2000. Leaf tissue turnover and efficiency of herbage utilization. En: Lemaire G, Hodgson J, De Moraes A, Nabinger C, Carvalho PC de F. [Eds.]. Grassland Ecophysiology and Grazing Ecology. Oxford : CABI Publishing. pp. 265 - 287.
Lemaire G, Chapman D. 1996. Tissue flows in grazed plant communities. En: Hodgson J, Illius AW. [Eds.].The Ecology and Management of Grazing Systems. Wallingford : CAB International. pp. 3 - 35.
Mannetje L´T. 2000. Measuring Biomass of Graslland Vegetation. En: Mannetje L´T, Jones RM [Eds.]. Field and laboratory methods for grassland and animal production research. New York : CABI Publishing pp. 151-177.
]]>Matthew C, Assuero SG, Black CK, Sackville Hamilton NR. 2000. Tiller dinamics of grazed swards. En: Lemaire G, Hodgson J, De Moraes A, Nabinger C, Carvalho PC de F. [Eds.]. Grassland Ecophysiology and Grazing Ecology. Oxford : CABI Publishing. pp. 127 - 150.
Nelson CJ, 2000. Shoot morphological plasticity of grasses: leaf growth vs. tillering. En: Lemaire G, Hodgson J, De Moraes A, Nabinger C, Carvalho PC de F. [Eds.]. Grassland ecophysiology and grazing ecology. Oxford : CABI Publishing. pp. 101 - 126.
Parsons AJ. 1988. The effects of season and management on the growth of grass swards. En: Jones MB, Lazenby A. [Eds.]. The grass crop : The physiological basis of production. London: Chapman and Hall. pp. 129 - 177.
Parsons AJ, Chapman DF. 2000. The principles of pasture growth and utilization. En: Hopkins A. [Ed.].Grass : its production and utilization. Oxford : Backwell Science. pp. 31 - 89.
Pérez MT, Hernández Garay A, Pérez J, Herrera JG, Bárcena R. 2002. Respuesta productiva y dinámica del rebrote de ballico perenne a diferentes alturas de corte. Técnica Pecuaria en México, 40: 251 - 263.
]]>Prache S, Peyraud J.1997. Préhensibilité de l´herbe pâturée chez les bovins et les ovins. INRA Productions Animales, 10: 337 - 390.
Rhodes I, Collins RP. 1993. Canopy structure. En: Davies A, Baker RD, Grant SA, Laidlaw AS. [Eds.]. Sward Measurement Handbook. Reading : The British Grassland Society. pp. 139 - 156.
Richards JH. 1993 Physiology of plants recovering from defoliation. En: Proceedings of the 17th International Grassland Congress. Wellington : SIR Publishing. pp. 85 - 94.
Saldanha S, Boggiano P, Cadenazzi M. 2010. Intensidad del pastoreo sobre la estructura de una pastura de Lolium perenne cv Horizon. Agrociencia, 14(1): 44 - 54.
Shaw NH, ´t Mannetje L, Jones RM, Jones RJ. 1985. Pasture Measurements. En: Shaw NH, Bryan WW. [Eds.]. Tropical pasture research : principles and methods. Hurley : CAB. pp. 235 - 250.
]]>Skinner RH, Nelson CJ. 1995. Elongation of the grass leaf and its relationship to the phylochron. Crop Science, 35: 4 - 10.
Skinner RH, Nelson CJ. 1994. Role of leaf appearance rate and the coleoptile tiller in regulating tiller production. Crop Science, 34: 71 - 75.
Tallowing JRB, Williams JHH, Large RV. 1986. Some consequences of imposing different continuous grazing pressures in the spring on sward morphology, herbage quality and the performance of young beef cattle. Journal of Agricultural Science, 106: 129 - 139.