Colnago Paula1, González Pablo2, Noguez Manuel1, Bentancur Oscar3, Galván Guillermo A.1
1Departamento de Producción Vegetal. Centro Regional Sur . Facultad de Agronomía, Universidad de la República. Camino Folle km 35,500 s/n. Progreso 90300, Canelones, Uruguay. Correo Electrónico: horticrs@fagro.edu.uy
2Departamento de Protección Vegetal. Facultad de Agronomía, Universidad de la República.
3Departamento de Biometría, Estadística y Cómputos. Estación Experimental Mario A. Cassinoni. Facultad de Agronomía, Universidad de la República
]]>
Recibido: 28/12/10 Aceptado: 2/9/11
Resumen
El mildiú causado por Peronospora destructor constituye una de las enfermedades de mayor importancia económica en la producción de cebolla. El objetivo de este trabajo fue identificar fuentes de resistencia en el germoplasma local de cebolla. Se realizaron ensayos a campo en 2007 y 2008 en los que se evaluaron en total 40 materiales genéticos. La incidencia de P. destructor se evaluó semanalmente, se ajustó una ecuación logística a los datos y se calculó el Área debajo de la Curva de Progreso de la Enfermedad (AUDPC). En 2007, el germoplasma local mostró diversidad en la incidencia de la enfermedad (3 a 99%), en la severidad (7 a 40% del área foliar afectada) y en el AUDPC. En 2008, los materiales evaluados también mostraron diferencias, aunque de menor magnitud. Se encontró relación entre el ciclo fenológico y la resistencia. El grupo de accesiones de día largo presentó mayor resistencia que los grupos de día intermedio y de día corto. Esto podría ser el resultado de factores de resistencia específicos presentes en el germoplasma de día largo, o de diferencias fisiológicas relacionadas al estado de desarrollo que condicionan la susceptibilidad al patógeno. En 2008, la evolución de la enfermedad fue muy variable. La incidencia de P. destructor cayó bruscamente en una evaluación, y subió a la semana siguiente hasta 100% de las plantas para varias accesiones. La diversidad observada en la respuesta a P. destructor confirma el potencial del germoplasma local para ser usado como fuente de resistencia en el mejoramiento de cebolla.
]]>Palabras clave: ALLIUM CEPA L., CICLO FENOLÓGICO, GERMOPLASMA LOCAL, PERONOSPORA DESTRUCTOR, RESISTENCIA PARCIAL
Summary
Downy Mildew (DM) caused by Peronospora destructor is one of the most economically important diseases in onion production. This research aimed to identify sources of resistance to DM in local onion germplasm. Field trials were conducted in 2007 and 2008 in order to evaluate a total of 40 genetic materials. The incidence of P. destructor was assessed weekly, a logistic equation was adjusted to the data, and the Area Under Disease Progress Curve (AUDPC) was calculated. In 2007, local germplasm showed differences in the proportion of diseased plants (3 to 99%), in severity (7 to 40% of total leaf area was affected) and AUDPC. In 2008, the materials tested showed differences as well, but smaller than in 2007. Resistance and phenological cycle were related. The group of long day accessions showed higher resistance than the groups with intermediate and short-day accessions. This behavior could involve specific resistance factors present in the germplasm of long day, or could be the result of physiological differences related to the stage of development that determine susceptibility to the pathogen. In 2008, evolution of disease was very variable. The incidence of P. destructor fell sharply on an assessment, and the following week rose to 100% of the plants for several accessions. Diversity observed in the response to P. destructor supports the potential value of local germplasm to be used as source of resistance in onion improvement.
Key words: ALLIUM CEPA L., PHENOLOGICAL CYCLE, LOCAL GERMPLASM, PERONOSPORA DESTRUCTOR, PARTIAL RESISTANCE
]]>
Introducción
La cebolla (Allium cepa L.) ocupa el tercer lugar por su Valor Bruto de Producción en la horticultura de Uruguay. La utilización de poblaciones locales que se mantienen mediante la multiplicación artesanal de semillas en los predios es alta. En 2004 el 74% de los productores utilizaron poblaciones locales de cebolla, ocupando el 50% del área para ese año (DIEA, 2008). Por otra parte, existen programas nacionales de mejoramiento genético de cebolla y de producción de semilla certificada. Unas 700 ha (35% del área total) son sembradas con semilla procedente de estos programas.
El mildiú de la cebolla causado por Peronospora destructor (Berk.) Casp. constituye una de las enfermedades de mayor importancia económica en la producción de cebolla en el mundo. En Uruguay actualmente es la enfermedad foliar de mayor importancia (González et al., 2011). Debido al uso incorrecto de los productos químicos comerciales disponibles para su control, no se logra un control exitoso, por lo que existe un alto uso de agrotóxicos con riesgos para el ambiente y para el agricultor sin alcanzar la eficacia deseada. En Uruguay se registran períodos de varias semanas al año con condiciones favorables para la epidemia, lo que hace difícil la implementación de estrategias de control exitosas (Maeso et al., 2000).
Peronospora destructor es un parásito obligado que pertenece al reino Chromista, phyllum Oomycota, clase Oomycete. Los síntomas característicos producidos por el patógeno son áreas ovales o cilíndricas de color verde-amarillo pálido a café, tanto en hojas como en escapos florales durante la etapa reproductiva (Schwartz y Mohan, 2008). Sobrevive en restos de cultivos, en bulbos y semillas infectadas (Schwartz y Mohan, 2008; Agrios, 2005; Maude, 1990).
]]>
Dentro de la base genética del cultivo no se dispone de fuentes de alta resistencia a P. destructor. Sin embargo, Allium roylei Stearn, una especie que pertenece a la sección Cepa del género Allium, presenta resistencia completa a P. destructor (Van der Meer y De Vries, 1990). La resistencia de A. roylei es independiente de la densidad del inóculo y de la edad de la planta (Kofoet y Zinkernagel, 1989). El cruzamiento con Allium cepa es viable, y el híbrido interespecífico resultante es resistente (Kofoet et al., 1990). Si bien se han logrado avances, existen barreras interespecíficas que dificultan la introgresión (Alan et al., 2003; Scholten et al., 2007), y solo en el largo plazo puede preverse la utilización de esta fuente de resistencia en cebollas adaptadas a diferentes regiones de producción.
En condiciones de producción en Uruguay se han observado diferencias dentro del germoplasma local y entre cultivares en la respuesta ante la enfermedad. La identificación de fuentes de resistencia parcial permitiría su explotación posterior en el mejoramiento genético. Estas diferencias en el desarrollo de la enfermedad podrían ser consecuencia de diferencias en componentes de la resistencia como la tasa de infección, el período de incubación y/o el período de latencia.
El desarrollo de cultivares con resistencia genética se presenta como una alternativa tendiente a reducir el uso de agrotóxicos, disminuyendo la posible contaminación del medio ambiente y protegiendo la salud humana. El objetivo de este trabajo fue caracterizar el germoplasma local de Uruguay en su respuesta a Peronospora destructor e identificar fuentes de resistencia. Se discuten los factores que podrían determinar las diferencias en el germoplasma local.
Se realizaron tres ensayos a campo, uno en 2007 y dos en 2008, que se ubicaron en el Centro Regional Sur (CRS), Facultad de Agronomía, Progreso, Canelones (Latitud Sur 34°36´49´´; Longitud Oeste 56°13´04´´).
Ensayo 2007
Se sembraron 27 poblaciones locales de Allium cepa y dos cultivares nacionales que se incluyeron como testigos: «Pantanoso del Sauce CRS» e «INIA Naqué» (Cuadro 1). Estas poblaciones abarcaron distintas regiones del país y tipos varietales: precoces (día corto, DC), semi-precoces (día intermedio, DI) y tardías (día largo, DL).
]]>El almácigo se sembró el 17 y 18 de abril de 2007 en canteros solarizados. El diseño experimental fue alfa-látice con cuatro repeticiones por accesión en bloques completos. El trasplante se realizó el 10 de agosto de 2007. Cada parcela contó con 60 plantas. Para aumentar y homogeneizar la presión de inóculo, cada dos canteros con las parcelas de evaluación, se intercaló una variedad susceptible (Pantanoso del Sauce CRS) a alta densidad. En el área donde se instaló el ensayo, se había cultivado cebolla durante los dos años anteriores, y hubo presencia de P. destructor. El control de malezas consistió en una aplicación de Oxifluorfen (1 l/ha) pre-trasplante y posteriormente carpidas manuales. A fines de octubre comenzó un período seco, por lo que el ensayo se regó para incrementar las condiciones predisponentes a la enfermedad. Las cosechas se realizaron a partir del 4 de diciembre hasta el 11 de enero. No se realizaron aplicaciones de fungicidas ni insecticidas.
Ensayos 2008
En un primer ensayo se sembraron diez poblaciones locales y los cultivares «INIA Casera» e «INIA FAGRO Dulce» (Cuadro 1). El diseño experimental fue de bloques completos aleatorizados con cuatro repeticiones por accesión. Cada parcela contó con 51 plantas y se intercaló el mismo cultivar susceptible. Los almácigos se sembraron el 23 de abril de 2008 y el trasplante se realizó el 11 de agosto de 2008. En el control de malezas se utilizó Linuron (1,5 kg/ha), y re-aplicaciones de Oxifluorfen + Oxadiazon a los caminos (0,5 l/ ha + 1,5 l/ ha). Posteriormente se complementó con carpidas manuales. No se realizaron aplicaciones de fungicidas ni insecticidas. Las cosechas se realizaron a partir del 27 de noviembre y se extendieron hasta el 12 de diciembre.
En el ensayo 2, se incluyeron dos poblaciones locales, el cultivar «Canarita CRS» y una población avanzada de mejoramiento denominada Endocría. La fecha de transplante, la densidad y el diseño experimental fueron los mismos que en el ensayo 1. Las cosechas comenzaron el 5 de diciembre y se extendieron hasta el 10 de enero de 2009.
]]>Los materiales que se evaluaron en 2008 fueron en su mayoría de DI. Cinco poblaciones locales (las accesiones UR9704, UR9706, UR9718, UR9720, UR9727) ya se habían evaluado en 2007. Para otras poblaciones locales ya evaluadas en 2007, no se contó con semilla suficiente para la instalación del ensayo 2008.
Evaluaciones realizadas
Las observaciones se realizaron sobre las plantas de la fila central de cada parcela. La incidencia (presencia/ausencia de manchas o signo) se midió semanalmente a partir del 27 de setiembre hasta el 23 de noviembre, y en 2008 desde el 4 de setiembre hasta el 31 de octubre. La severidad se registró el 22 de noviembre en 2007 y el 3, 10 y 17 de octubre en los ensayos de 2008. Con los valores de incidencia de la enfermedad se calculó el área bajo la curva de progreso de la enfermedad (AUDPC). Para la evaluación de severidad se estimó el porcentaje del área foliar afectada por la enfermedad, utilizando una escala de 0 a 100% con intervalos de 5% (Agrios, 2005). La evaluación se realizó sobre hojas verdes activas, y se tuvo en cuenta la presencia de esporulación o síntomas típicos de P. destructor. No se consideraron hojas completamente secas o senescentes ni las puntas secas de hojas que no permitían reconocer si la causante de la senescencia del tejido fue P. destructor, otra enfermedad u otra causa.
]]>
Análisis estadístico
En 2007, el efecto de las accesiones sobre la incidencia fue estudiado mediante el ajuste de regresiones logísticas en función del tiempo. Se asumió una distribución binomial de plantas afectadas en el total de plantas por población. Las curvas de incidencia se compararon mediante contrastes de razón de verosimilitud (LRT, likelihood ratio test). También se efectuaron contrastes entre grupos de accesiones fenológicos, determinados de acuerdo al ciclo (momento de cosecha). Se utilizó el procedimiento GENMOD del paquete estadístico SAS versión 9.1.3 (SAS Institute, Cary, NC). Para los datos del ensayo 2008 no se ajustaron curvas, sino que directamente se estimó la probabilidad de incidencia en cada fecha de muestreo. Las probabilidades de incidencia de las diferentes poblaciones y momentos se compararon usando el test de Tukey adaptado por Kramer (1956).
El efecto de las poblaciones sobre la severidad en cada año fue estudiado ajustando modelos lineales generalizados asumiendo una distribución multinomial ordinal de la escala de severidad. En 2008 se consideró el efecto del momento de evaluación en los modelos. Se usó también el procedimiento GENMOD del mismo paquete estadístico.
Resultados
Ensayo 2007
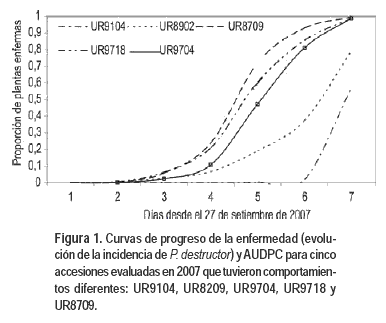
El germoplasma local de cebolla mostró diversidad en la incidencia y severidad (Cuadro 2, Figura 1). Si bien todas las poblaciones se comportaron como susceptibles, el progreso de la incidencia no fue similar para las diferentes accesiones (p < 0,05). La evaluación del 9 de noviembre fue la fecha en que más difirieron las poblaciones, en un rango de 3 a 99% de plantas enfermas. A partir de entonces se observó un rápido progreso de la enfermedad para todos los materiales genéticos. Las accesiones UR9104 y UR8902 fueron las que tuvieron menor porcentaje de plantas enfermas (Cuadro 1). En el otro extremo, las accesiones que tuvieron mayor incidencia fueron UR9718, UR8709 y UR9704 con 92, 95 y 99% de incidencia respectivamente el 9 de noviembre. Como consecuencia, las accesiones también difirieron fuertemente en el área bajo la curva de progreso de la enfermedad (Figura 1).
]]>
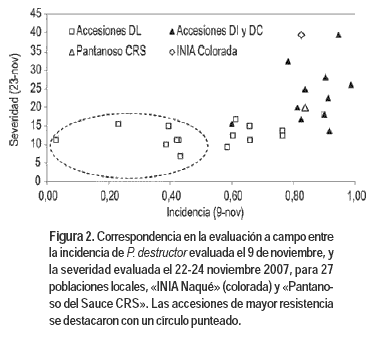
La severidad a P. destructor fue evaluada solo el 22-24 noviembre de 2007. Se contó con una sola fecha de monitoreo ya que las condiciones ambientales favorables al desarrollo del patógeno no se extendieron por un período prolongado. Se observó diversidad en un rango desde 7 a 40% del área foliar afectada (Cuadro 1). Muchas de las accesiones que mostraron menor incidencia el 9 de noviembre fueron las mismas que presentaron menor porcentaje de tejido afectado el 23 de noviembre: UR8902 con severidad estimada en 6,9% y UR9104 con 11,3%. Por lo tanto, al combinar el comportamiento en cuanto a incidencia y severidad de P. destructor, pudo diferenciarse un grupo de accesiones que presentó los menores valores para ambas variables (Figura 2): valores inferiores a 45% en la incidencia, y menor o igual a 20% en la severidad. Este grupo quedó integrado por las accesiones UR9104, UR8902, UR8815, UR8701, UR8703, UR8906 y UR8823.
]]> Los cultivares «INIA Naqué» y «Pantanoso del Sauce CRS» incluidos en el ensayo como testigos, se ubicaron dentro del grupo de materiales genéticos más susceptibles (Cuadro 1, Figura 2).
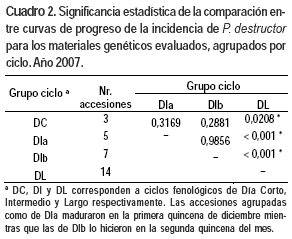
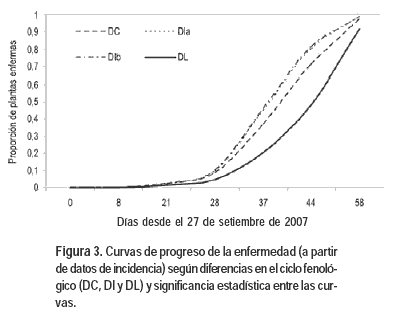
Al analizar la relación de la respuesta a P. destructor con variables morfológicas y agronómicas, se encontró que estuvo relacionada con la variación en el ciclo fenológico (Figura 3). Es así que los materiales genéticos tempranos (DC y DI) fueron significativamente más susceptibles que materiales tardíos (DL) (Cuadro 2). Si bien las diferencias observadas entre grupos fenológicos son de mayor magnitud, también se observó diversidad en la susceptibilidad a P. destructor dentro de los grupos fenológicos de DI y DL (Cuadro 3).

]]>
Ensayos 2008
En 2008 se observaron síntomas a partir del 4 de setiembre, a diferencia de lo observado para 2007, donde se registraron los primeros síntomas un mes más tarde, a partir del 4 de octubre. En ambos años se llegó a valores de incidencia de la enfermedad del 100% para los cultivares y poblaciones más susceptibles. En 2007 estos valores se registraron el 23 de noviembre, con un rango que osciló entre 65% y 100% de incidencia de la enfermedad. En 2008 la incidencia máxima se alcanzó el 17 de octubre y el rango fue de 80% a 100%.
En esta temporada no se pudo ajustar un modelo matemático para el progreso de la enfermedad, por lo que se analizaron las medias de cada población en cada fecha. En la evaluación de la enfermedad no se consideraron las hojas secas, lo que pudo haber distorsionado el registro de la enfermedad. Se observó una caída brusca en el registro de incidencia y severidad (10 de octubre) y posteriormente valores altos alcanzando el 100% de incidencia (17 de octubre) en la mayoría de las accesiones al darse condiciones para nuevos ciclos de infección.
]]>
No hubo diferencias significativas por fecha de monitoreo, pero sí se observaron diferencias en el comportamiento general de las poblaciones (Cuadro 4). En el ensayo 1 se destacó UR9719 por tener menor incidencia a lo largo del período estudiado. En el ensayo 2, el cultivar «Canarita CRS» fue el que presentó los menores valores promedio de incidencia para el período (Cuadro 4). Dado que las accesiones corresponden en su mayoría al tipo DI, las diferencias en la respuesta a la enfermedad observadas entre accesiones no son dependientes del ciclo.
En el ensayo 1, las accesiones UR9719 y UR9706 fueron las accesiones con menor severidad, ya que no superaron el 26,5% de tejido afectado. Por el contrario, UR9704 y el cultivar «INIA Casera» fueron los que presentaron peor comportamiento, así como la mayor variabilidad en los datos en las tres fechas consideradas. La magnitud de las diferencias en severidad, sin embargo, fue menor que las diferencias observadas para incidencia de la enfermedad a lo largo de las distintas fechas de monitoreo (Figura 4). En el ensayo 2, «Canarita CRS» presentó los menores valores de tejido afectado y menor variabilidad en la distribución de los datos (Figura 4).
Entre las accesiones que presentaron menor incidencia de P. destructor (UR9719 y UR9706 en el ensayo 1, y «Canarita CRS» en el ensayo 2), también la severidad fue menor, resultando en menor porcentaje de tejido afectado.
El análisis de la información climática para el período de evaluación de las accesiones mostró variaciones importantes en cuanto a las condiciones ambientales predisponentes a la enfermedad. Se analizaron los días y la duración del período (cantidad de horas en esos días) en los que se dieron las condiciones para esporulación de P. destructor (oscuridad, humedad relativa igual o mayor a 95%, y temperatura en el rango de 4 a 23 °C).
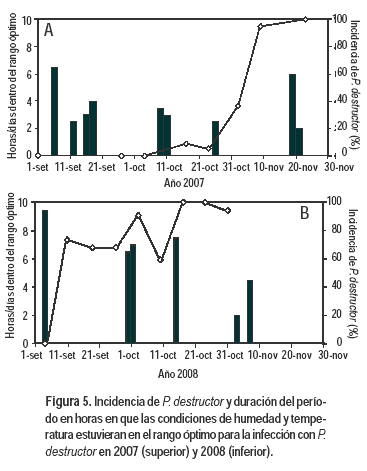
En 2008 se observó que las oscilaciones de temperatura y humedad fueron mayores que en 2007 para el período estudiado (Figura 5). En efecto, los síntomas de la enfermedad comenzaron más temprano en la estación de crecimiento, aunque posteriormente se dieron lapsos prolongados sin condiciones para la esporulación, o períodos con condiciones óptimas discontinuos. Esto coincide con las fuertes oscilaciones en el progreso de la enfermedad observadas en 2008, tanto en la incidencia como en la severidad. Al mismo tiempo, en este segundo año, en los períodos en que se dieron condiciones para infección y o esporulación, esas condiciones persistieron por mayor tiempo que el año anterior (en 2007 no superan en ningún caso las 6,5 horas).
Tanto en 2007 como en 2008 se observó que el progreso de la enfermedad estuvo más asociado a lo que se definió como el rango óptimo de temperaturas (10 a 14 ºC), y no al rango amplio de ocurrencia de la enfermedad (4 a 23 ºC). Por ejemplo, en 2007 los primeros registros de incidencia de la enfermedad fueron en torno al 15 de octubre, y podrían ser consecuencia de condiciones óptimas que se registraron los días 9 y 11 de octubre. Estas condiciones se volvieron a producir el 29 de octubre causando un aumento del número de plantas que presentaron síntomas activos el 4 de noviembre. En 2008, los primeros síntomas se observaron el 14 de setiembre, luego de días con condiciones óptimas los primeros días de ese mes (Figura 5).
]]> 
Las accesiones UR9704, UR9706, UR9718, UR9720 y UR9727 se evaluaron en las zafras 2007 y 2008. Los resultados obtenidos en el comportamiento sanitario de estos materiales son consistentes entre zafras. La accesión UR9706 fue una de las destacadas dentro del grupo fenológico de DI en 2007 por presentar baja incidencia y severidad a P. destructor, lo que coincide con los resultados obtenidos en las evaluaciones de 2008. Entre estas accesiones, UR9704 fue la que presentó mayor número de plantas enfermas y de tejido afectado en ambos años (Cuadros 1 y 4).
Discusión
Diferentes mecanismos involucrados en la resistencia parcial pueden ocasionar un atraso en la instalación del patógeno así como una tasa de progreso de la enfermedad menor (Agrios, 2005; Vanderplank, 1984), como se observó con las accesiones UR9104 y UR8902 en el ensayo de 2007, así como con UR9119 y el cultivar «Canarita CRS» en los ensayos de 2008.
Una posible explicación de las diferencias observadas entre las poblaciones locales puede involucrar las defensas estructurales de la planta. A este nivel podrían estar actuando elementos constitutivos como la capa de cera que recubre toda la superficie foliar, presenta variación genética entre cultivares y accesiones y tiende a disminuir la probabilidad de que se forme una película de agua continua en la superficie foliar, elemento que es esencial para la infección de P. destructor (Schwartz y Mohan, 2008; Hildebrand y Sutton, 1982). Esta característica es importante en el caso del patosistema A. cepa « Botrytis squamosa (Galván et al., 2004). El ángulo de inserción de las hojas en el falso tallo que lleve a diferencias en la persistencia de las gotas de lluvia o rocío sobre la superficie foliar, es otro parámetro que puede estar influyendo en las diferencias observadas entre accesiones y cultivares (Kofoet y Zinkernagel, 1989).
Otra hipótesis que explicaría las diferencias en la incidencia de P. destructor entre accesiones son las diferencias en las respuestas desencadenadas ante la presencia del patógeno por el sistema de defensa de la planta, generando diferencias en la tasa de desarrollo de la enfermedad. Diferencias bioquímicas y estructurales a nivel histológico una vez producida la reacción de reconocimiento pueden determinar qué tan rápido el patógeno se desarrollará, y qué tan severos serán los síntomas.
]]>Esto puede evidenciarse a través de diferencias en el período de latencia así como en el número de infecciones exitosas (Niks et al., 2011). Al trabajar en condiciones de campo, las diferencias que se observaron en cantidad total de enfermedad a través de AUDPC en 2007 o en el momento en que se llega al mayor porcentaje de plantas afectadas en 2008, son producto de varios ciclos de infección, esporulación y dispersión del patógeno.
Relación con el ciclo fenológico
En el ensayo de 2007, aun cuando accesiones de DC, DI y de DL fueron trasplantadas en la misma fecha, las accesiones de DL fueron las menos afectadas, por lo que la diferencia en la respuesta a P. destructor no se debe a un escape por época. Una posible hipótesis que explique esta relación con el ciclo fenológico es que el comportamiento frente a P. destructor podría atribuirse a factores de resistencia específicos en el grupo de materiales tardíos (valencianas), de expresión cuantitativa (Kamoun et al., 1998; Vanderplank, 1984). De ser así, podría transferirse la resistencia a los otros grupos fenológicos del germoplasma de cebolla mediante cruzamientos y selección recurrente por resistencia.
]]>
Alternativamente, otra hipótesis podría ser que las diferencias en la respuesta a P. destructor son resultado de diferencias entre los grupos de madurez según estado fisiológico-fenológico. En accesiones de DC o DI para las que se ha iniciado la bulbificación o están próximos a la bulbificación en el momento en que aparece P. destructor, podrían producirse cambios hormonales y nutricionales que afecten negativamente mecanismos de resistencia basales. En ese caso, la resistencia parcial en las accesiones de día largo no sería fácilmente aplicable al desarrollo de cultivares precoces y semiprecoces resistentes.
Esta segunda hipótesis vincula la expresión de resistencia con el desarrollo del cultivo. La adquisición de resistencia en etapas tardías del desarrollo (resistencia de la planta adulta), es un fenómeno comúnmente observado para diversos cultivos y pato-sistemas. La interacción P. destructor-A. cepa parece manifestar lo opuesto: hay mayor susceptibilidad conforme avanza su desarrollo. Los mecanismos involucrados en la «resistencia relacionada a la edad» (RRA), denominación propuesta por Lazarovits et al. (1981), difieren de otros relativos al sistema de defensa innato, la resistencia sistémica inducida o la resistencia sistémica adquirida. La resistencia a enfermedades en los casos de RRA está asociada a cambios de fase o transiciones importantes ocurridas a lo largo del ciclo de vida de la planta (Poethig, 2003; Bäurle y Dean, 2006). En el ensayo de 2007 se observó que las accesiones correspondientes a DL tuvieron mayor resistencia que las de DI y las de DC. Dada la respuesta cualitativa al fotoperíodo que tiene esta especie para bulbificar, aun cuando todas las accesiones fueron plantadas al mismo tiempo, el estado de desarrollo en el que se encontraban al momento que comenzó la infección, difería fuertemente entre grupos. Los requerimientos de fotoperíodo se cumplieron en torno al 1-7 de octubre, al 1-7 de noviembre, y al 5-10 de diciembre, para los ciclos de DC, DI y DL respectivamente. En la evaluación del 4 de octubre, sólo 3 de las 15 accesiones de DL presentaban síntomas, 4 de las 11 accesiones de DI y 2 de las 3 accesiones de DC. En la evaluación del 2 de noviembre, estos valores ascienden a 14 accesiones de DL, y la totalidad de las accesiones de DI y de DC, pero los valores de incidencia difirieron significativamente entre grupos fenológicos. Se encontraron los menores valores para DL con un promedio de 21% de plantas enfermas, mientras que el promedio para DI y DC fue de 34% y 35% respectivamente. En síntesis, conforme avanza el desarrollo fenológico, la planta se hace más susceptible, pero no puede asociarse claramente un cambio de fase como determinante sino que parecería que a partir del inicio de la bulbificación la planta está más susceptible en términos cuantitativos. Sumado a esto, al tratarse de un patógeno policíclico, lo observado podría ser el resultado de varios ciclos que determinan un rápido crecimiento de la enfermedad, en la medida de que las condiciones ambientales se mantengan dentro de los rangos óptimos.
Los mecanismos que controlan la transición del desarrollo en la planta huésped pueden ser los mismos que gobiernan la expresión de la resistencia (Pierre et al., 2007). El gran número de genes que interviene en la resistencia parcial, podría estar vinculado a las distintas etapas de los procesos fisiológicos de la planta, que generan las sustancias y estructuras que constituyen sus mecanismos de defensa (Agrios, 2005).
Temperatura y humedad son los principales factores climáticos que afectan el desarrollo de la epidemia. La temperatura y humedad relativa por fuera del intervalo óptimo para el desarrollo del patógeno resultaron en un descenso importante de la enfermedad registrada en la semana del 3 al 10 de octubre en 2008. Estas condiciones podrían estar afectando negativamente su sobrevivencia (cantidad de inóculo que permanece viable), dispersión y/o su tasa de reproducción. El desfasaje entre (1) el momento del ciclo en el que se dieron las condiciones óptimas para la expresión de la enfermedad con (2) el momento en que se registraron aumentos en la incidencia y/o severidad, puede estar explicado por el período de latencia, si bien no puede ser determinado con precisión en condiciones de campo.
El modelo logístico que se ajustó para 2007 tiene una serie de supuestos. Entre ellos, que el ambiente en el cual la enfermedad se desarrolla es uniforme y que hay cambios constantes en la enfermedad que son inmediatamente visibles (Campbell, 1998). Dadas las mayores oscilaciones climáticas durante la evaluación de 2008, el incumplimiento de estos dos supuestos posiblemente sea lo que haya imposibilitado el ajuste de una curva logística. No obstante, no es posible determinar una fuerte asociación entre la evaluación preliminar de las condiciones requeridas para la enfermedad y las curvas construidas tras las evaluaciones de 2007 y 2008. Este análisis debería realizarse considerando el período de latencia del patógeno y con mayor conocimiento acerca de cómo las fluctuaciones climáticas actúan sobre su ciclo.
La caída brusca en la incidencia (plantas registradas como enfermas en mediciones anteriores pasaron a contabilizarse como sanas) podría explicarse por la senescencia total del tejido que estuvo enfermo, y por la recuperación de áreas foliares. El crecimiento del hospedero es un factor importante en muchas epidemias y puede afectar la tasa aparente de crecimiento de la enfermedad y la pendiente de la curva de progreso de la enfermedad (Campbell, 1998). El crecimiento de las plantas puede «diluir» el efecto de la enfermedad. Si la planta crece a mayor velocidad que lo que lo hace el patógeno es posible incluso observar que «decrece» la enfermedad aun cuando no lo esté haciendo. Un factor adicional es la pérdida de hojas viejas de la planta y/o de hojas enfermas, lo que puede causar que la cantidad de enfermedad observada disminuya en el tiempo. Ambos factores, crecimiento y defoliación pueden ser importantes en el intento de calcular la tasa de crecimiento de la enfermedad, pudiendo ser subestimada o incluso negativa (Campbell, 1998).
]]>Se han propuesto distintas correcciones para poder utilizar estos modelos en simulación de epidemias (Campbell, 1998; Rabbinge et al., 1989; Vanderplank, 1984; Kushalappa y Ludwig, 1982). Estas correcciones proponen distintas formas de considerar la enfermedad referida al crecimiento del hospedero, que no pudieron aplicarse en este trabajo, en la medida que no se evaluaron variables de crecimiento.
Conclusiones
Las diferencias observadas en incidencia y severidad representan un elemento valioso para el control de la enfermedad, en particular en un esquema de control integrado. La resistencia parcial con que cuentan algunos de los materiales evaluados, por ejemplo UR9104 y UR8902, no evita que la planta se enferme sino que retarda el desarrollo de la infección y por tanto la propagación de la enfermedad y el desarrollo de la epidemia.
]]>
Si bien se requiere de más años de evaluación, los resultados son promisorios en cuanto a la utilización de estas accesiones como fuentes de resistencia parcial en programas de mejoramiento.
Agradecimientos
Al Dr. Diego Maeso, por aportes realizados durante el monitoreo de la enfermedad, y sugerencias sobre el manuscrito de este trabajo. A los funcionarios de horticultura del CRS, Técn. Agr. V. Ferreira, N. Curbelo, y los Sres. N. Jara y O. Costa. Este trabajo se realizó como parte del proyecto FPTA-INIA 250 «Resistencia a enfermedades en cebolla».
Agrios GN. 2005. Plant Pathology. Florida: Elsevier Academic Press. 921p.
Alan AR, Mutschler MA, Brants A, Cobb E, Earle ED. 2003. Production of gynogenic plants from hybrids of Allium cepa L. and A. roylei Stearn. Plant Science, 165 (6): 1201- 1211.
Bäurle I, Dean C. 2006. The timing of developmental transitions in plants. Cell, 125: 655 - 664.
Campbell CL. 1998. Disease progress in time: modelling and data analysis. En: Jones G. [Ed.]. The epidemiology of plant diseases. Netherlands : Springer. pp. 42 - 72.
DIEA. 2008. Encuestas hortícolas 2007 : Zonas Sur y Litoral Norte. Montevideo : MGAP. 44p. (Serie Encuestas ; 263).
Galván G, González P, Reggio A. 2004. Onion leaf blight caused by Botrytis squamosa in Uruguay and the differential response of local cultivars. Allium Improvement Newsletter, 13: 51 - 54.
González PH, Colnago P, Peluffo S, González Idiarte H, Zipitria J, Galván G.A. 2011. Quantitative studies on downy mildew (Peronospora destructor Berk. Casp.) affecting onion seed production in southern Uruguay. European Journal of Plant Pathology, 129: 303 - 314.
Hildebrand PD, Sutton JC. 1982. Weather variables in relation to an epidemic of onion downy Mildew. Phytopathology, 72(2): 219 - 224.
Kamoun S, Van West P, Govers F. 1998. Quantification of late blight resistance of potato using transgenic Phytophthora infestans expressing B-glucuronidase. European Journal of Plant Pathology, 104: 521 - 525.
Kofoet A, Kik C, Wietsma WA, De Vries JN. 1990. Inheritance of resistance to downy mildew (Peronospora destructor (Berk.) Casp.) from Allium roylei Stearn in the backcross Allium cepa L. x (A. Roylei x A. cepa). Plant Breeding, 105: 144 - 149.
Kofoet A, Zinkernagel V. 1989. Resistance to downy mildew (Peronospora destructor (Berk.) Casp.) in Allium species. Journal of Plant Disease and Protection, 97(1): 13 -23.
Kramer CY. 1956. Extension of multiple range tests to group means with unequal numbers of replications. Biometrics, 12: 309 - 310.
Kushalappa AC, Ludwig A. 1982. Calculation of apparent infection rate in plant diseases: Development of a method to correct for host growth. Phytopathology, 72: 373 - 377.
Lazarovits G, Stössel R, Ward EWB. 1981. Age-related changes in specificity and glyceollin production in the hypocotyl reaction of soybeans to Phytophthora megasperma var. sojae. Phytopathology, 71: 94 - 97.
Maeso D, Arboleya J, Fernández S, Suárez C, Medina V. 2000. Evaluación de un sistema de pronóstico para el control de enfermedades foliares en diferentes cultivares de cebolla. En: Presentación de resultados experimentales en ajo y cebolla: Reunión de divulgación. Montevideo: INIA. (Actividades de difusión; 223). pp. 17 - 29.
Maude RB. 1990. Leaf diseases of onions. En: Rabinowitch HD, Brewster JL. [Eds.]. Onions and Allied crops. Vol. 2. Florida: CRC Press. pp. 173 - 190.
Niks RE, Parlevliet JE, Lindhout P, Bai Y. 2011. Breeding crops with resistance to diseases and pests. Enfield : Enfield Pub & Distribution Co. 200p.
Pierre M, Riviere D, Galiana E. 2007. Resistance to pathogens and host developmental stage: a multifaceted relationship within the plant kingdom. New Phytologist, 175: 405 - 416.
Poethig RS. 2003. Phase change and the regulation of developmental timing in plants. Science, 301: 334 - 336.
Rabbinge R, Zadoks JC, Bastiaans L. 1989. Population models. En: Rabbinge R, Ward SA, van Laar HH. [Eds.]. Simulation and systems management in crop protection. Wageningen : Pudoc. pp. 81 - 118.
Scholten OE, Van Heusden AW, Khrustaleva LI, Burger-Meijer K, Mank RA, Antonise RGC, Harrewijn JL, Van Haecke W, Oost EH, Peters RJ, Kik C. 2007. The long and winding road leading to the successful introgression of downy mildew resistance into onion. Euphytica, 156: 345 - 353.
Schwartz HF, Mohan SK. 2008. Compendium of onion and garlic diseases. St Paul: APS Press. 54p.
Van der Meer QP, De Vries JN. 1990. An interspecific cross between Allium roylei Stearn and Allium cepa L., and its backcross to A. cepa. Euphytica, 47: 29 - 31.
Vanderplank JE. 1984. Disease resistance in plants. Orlando: Academic Press. 194p.