











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references

 Permalink
Permalink
1. Introduction
The eradication of a pest raises questions about the species role in the environments it inhabits, its ecological interactions network and, ultimately, what harmful effects the control program could cause on the ecosystems. Currently, the eradication of the New World Screwworm (NWS) fly1, Cochliomyia hominivorax (Diptera: Calliphoridae), an obligatory ectoparasite that causes myiasis in warm-blooded vertebrates, including humans2, is being discussed in Uruguay. The economic losses due to myiasis for the Uruguayan livestock sector were evaluated through different strategies, and estimated as USD 40-45 million per year1. In addition, more than 800 human cases annually, mainly among the rural population, are projected based on epidemiological data from the National Livestock Services (DGSG, MGAP)3)(4)(5. However, an underreporting of human cases is estimated due to the myiasis stigmatizing effect.
The NWS has been eradicated from North and Central America through a Wide Area Integrated Pest Management (WA-IPM) program that integrated the Sterile Insect Technique as its main tool, since 1957, when the control program began in the USA6)(7)(8. Since 2004, a permanent barrier to releasing of sterile insects has been maintained in the Darién region, on the border between Panama and Colombia, to prevent its reintroduction7. To our knowledge, no reports of negative impacts of NWS eradication programs on ecosystems or other species have ever been published.
Living organisms interact within the species level and between species in their community through complex networks. The loss of a species in a community can lead to secondary extinctions or cascading effects through its ecological interactions9)(10. The role of the lost species and the capacity of the remaining species with which it interacts to compensate for its loss are essential for the ecosystem functioning, determining the magnitude of the secondary effects in the ecological community10)(11)(12.
A preliminary risk analysis of the environmental impact of an NWS fly eradication program in Uruguay indicated that it would not result in significant alterations on ecosystems and ecological interactions1. As a complement, in this paper we describe the ecological interactions network that the NWS integrates, and assess whether secondary extinctions or cascading effects are likely to occur as a consequence of NWS eradication in Uruguay. Here, we summarize the main results related to the ecological impact of the eradication of the NWS obtained in a consultancy contracted by the IPA (Instituto Plan Agropecuario) and the MGAP (Ministry of Livestock), carried out during 2021 for the social and environmental impact assessment of the eradication program.
2. Ecological interactions
Scientific literature research was carried out to identify the direct ecological interactions of the NWS fly, which yielded a total of 274 documents, of which 79 were included here due to their relevance in describing the main ecological interactions (see the complete list of documents in Supplementary Material).
2.1 Parasitism
The NWS fly is an obligatory ectoparasite of warm-blooded vertebrates because it feeds on living tissues (i. e. myiasis) in its larval stages. The NWS develops its life cycle in an extensive list of hosts, including humans3)(4)(5)(6. Domestic hosts include all animals of production and companion2. Myiasis is more frequently described among wild hosts in zoos, and even today, there are few reports in wild animals13. Some of the wild animals in which myiasis has been recorded are the Texas opossum (Didelphis virginiana texensis (Didelphimorphia: Didelphidae))14, the water buffalo (Bubalus bubalis (Artiodactyla: Bovidae))15, the white-tailed deer (Odocoileus virginianus (Artiodactyla: Cervidae))14)(16, the key pygmy deer (O. virginianus clavium)17, the pampas deer (Ozotoceros bezoarticus (Artiodactyla: Cervidae))18, the fallow deer (Dama dama (Artiodactyla: Cervidae))17, the wild boar (Sus scrofa (Artiodactyla: Suidae))19-21, the capybara (Hydrochoerus hydrochaeris (Rodentia: Caviidae))22, the maned wolf (Chrysocyon brachyurus (Carnivora: Canidae))13, the ocelot (Leopardus pardalis (Carnivora: Felidae))23, the jaguar (Panthera onca (Carnivora: Felidae))24, the lesser grison (Galictis cuja (Carnivora: Mustelidae))25, and the rhea (Rhea americana (Struthioniformes: Rheidae))18.
Also, C. hominivorax parasites have been described, such as the Trichotrombidium muscarum mite (Acari: Prostigmata), mainly affecting adults26. This parasite feeds on the hemolymph of its hosts and probably on their eggs27. The micro-hymenoptera parasitoid wasps Aphaereta laeviuscula (Hymenoptera: Braconidae) and Nasonia vitripennis (Hymenoptera: Pteromalidae) were recorded parasitizing C. hominivorax pupae28. In addition, the hymenopterans wasps of the Superfamily Chalcidoidea (Insecta: Hymenoptera) were also identified in its pupae29. Finally, the fungus Aspergillus flavus (Fungi: Ascomycota) has been detected in C. hominivorax pupae29.
2.2 Predation
In its adult phase, some identified predators of C. hominivorax are the spiders Nephila clavipes, Eriophora ravilla, Neoscona oaxacensis (Araneae: Araneidae), Leucauge spp. (Araneae: Tetragnathidae)30, and spiders of the Family Zoridae (Araneae: Lycosoidea)29. Nephila clavipes has been recorded in Uruguay31. Others species were identified as predators of immature phases of C. hominivorax life cycle, such as Chrysomya albiceps (Diptera: Calliphoridae)32. However, the possible control that it can exert over C. hominivorax is low since it also can damage intact tissues, and its association causes more harm than benefits32. The ant species Labidus sp., Nomamyrmex sp., Solenopsis sp. and Dolichoderus sp. (Insecta: Hymenoptera) may attack C. hominivorax larvae once they leave the host29)(33. Also, most beetles of the Family Staphylinidae are predators of living carrion insects34. Usually, animals with myiasis hide for several days before death, and larvae wander between 0.5 and no more than 2 meters from where they land. If they mature on the dead, they are primarily found on the ground below the carcass33. The pupae, therefore, tend to appear in aggregations associated with those of carrion flies. Likely many mortality factors operate against prepupae and pupae in soils, including predation by beetles and other insects; however, they play a minor role in limiting the number of flies that develop around carrion33. According to Little35 larvae of several species of this Family (Staphylinidae) living in sympatry with adults are predators of pupae and larvae of Diptera, while others live in colonies of ants and termites. Most species feed on Diptera larvae in their adult phase, on tetrapod corpses. In general, they are considered beneficial insects34.
2.3 Commensalism
The NWS fly presents commensalism interactions, in some cases as commensal, and in others in the role of host. As a commensal, it interacts with species that generate skin or mucosal lesions, attracting and facilitating the oviposition and development of C. hominivorax larvae. Most important species are ticks (Arachnida: Ixodidae), such as Rhipicephalus microplus36, Rhipicephalus sanguineus37, Amblyomma maculatum14)(38)(39, A. aureolatum, A. dubitatum, A. tigrinum and A. triste37)(39)(40, and Ixodes aragaoi37)(41. However, A. aureolatum, A. dubitatum, A. tigrinum, A. triste and I. aragaoi do not usually cause severe enough lesions that facilitate the infestation by C. hominivorax larvae (2021 conversation with Tatiana Saporiti; unreferenced). Other lesion-causing species that facilitate NWS infestation are the head louse (Pediculus humanus capitis (Phthiraptera: Pediculidae))42, the human botfly (Dermatobia hominis (Diptera: Oestridae))42)(43)(44 and the ship botfly (Oestrus ovis (Diptera: Oestridae))45. Although C. hominivorax and D. hominis are primary myiasis-causing flies, they do not compete for resources, since their oviposition strategies are highly different.
On the other hand, the NWS fly also interacts with blowflies (Diptera: Calliphoridae) such as Cochliomyia macellaria46)(47)(48)(49, Chrysomya albiceps32)(42, Chrysomya rufifacies50, Lucilia cuprina51)(52)(53, Lucilia eximia51)(52 and Lucilia sericata51)(54. These other species of Diptera are fundamentally necrophagous, and when present in myiasis, they feed on the necrosed tissues generated. In northern Uruguay, 70.8% of myiasis showed other species of Diptera55.
Finally, in Libya in 1991, C. hominivorax adults were found infested with the cosmopolitan phoretic mite Macrocheles muscaedomesticae. This mite does not have an affinity for any particular host. It is likely that during the SIT program in Libya, the exposure to M. muscaedomesticae occurred due to the feeding behavior of the NWS fly. Calliphoridae feeds on animal feces to reach ovary maturation when carrion is not available27.
2.4 Mutualism
An often unrecognized but crucial ecological role of some Calliphoridae is pollination56)(57)(58)(59)(60)(61. The NWS fly adults feed on flower nectar, which is vitally important for their survival, ovary maturation and reproductive success, due to which they could play a role as pollinators60. However, abundance studies carried out in various regions using different types of rotten animal and plant baits indicated that the NWS has very low abundances concerning other nectarivorous Calliphoridae, such as C. macellaria and Chrysomya sp.62)(63)(64)(65, which is expected due to the high resources' availability for necrophagous calliphorids and the bias introduced by the baited traps used. Despite of that it should be noted that rotten liver baits are commonly used in surveys of NWS adults14)(66. Therefore, the overall contribution of the NWS fly as a pollinator is low relative to that of the other calliphorids with which it cohabits.
Facilitation between Diptera that causes obligatory myiasis is another mutualistic interaction. Wound infestation by NWS larvae does not exclude other species; on the contrary, it highly facilitates its presence and vice versa. Several myiasis-causing species inhabit in Uruguay, such as Chrysomya megacephala, Chrysomya rufifacies and Lucilia sericata67. But as the parasitism of these species is facultative and, instead, they develop mainly on carrion, the benefit of the interaction with C. hominivorax as a substrate provider is negligible.
2.5 Competition
No competitive interactions were identified between the NWS fly adult phase and other species, since it is a generalist (i. e., nectar, pollen and floral molasses), and food is not a limiting factor. Also, as mentioned before, the NWS fly myiasis does not generate competitive interactions.
In order to investigate the substitution or the increase of myiasis cases by other Calliphoridae after the eradication of C. hominivorax in Panama, larvae found in myiasis cases were taxonomically classified during the first years of the eradication program (1998-2005)68. Six species were identified: Dermatobia hominis (58%), Lucilia spp. (20%), C. macellaria (19%), C. rufifacies (0.4%), as well as larvae of Sarcophagidae (3%) and Muscidae (0.3%). The authors concluded that the absence of C. hominivorax did not increase myiasis caused by other species, supporting the hypothesis that there are no strong competitive interactions between C. hominivorax and the other species in Panama.
3. Impact of the eradication of the NWS fly on biodiversity
The gradual reduction of NWS fly populations until its eradication in Uruguay can impact the species and ecosystems on the national scale. However, in regions where the Sterile Insect Technique has been applied to control the NWS fly, local eradication usually occurs in less than one year. Nevertheless, it usually takes several years to achieve eradication in the entire region.
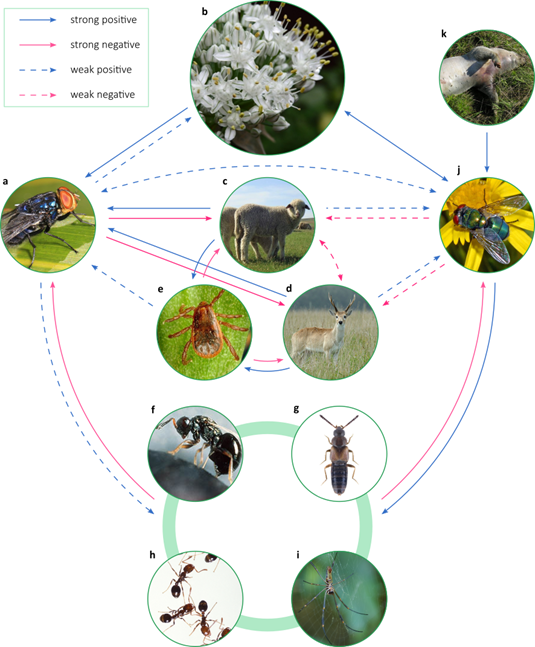
Figure 1: Conceptual synthesis of the NWS fly ecological interactions network obtained from literature review. Strong effects are those that may play an important role in regulating the interaction partner of a species. The images contain representative examples of assemblages of species that interact with the NWS fly. a: Cochliomyia hominivorax; b: pollinated plants (Allium cepa); c: domestic hosts (Ovis orientalis aries); d: wild hosts (Ozotoceros bezoarticus); e: ectoparasites that facilitate myiasis (Rhipicephalus sanguineus); f-i: parasitoids and predators; f: parasitoid microhymenoptera (Nasonia vitripennis); g: predatory staphylinid beetles (Atheta coriaria); h: predatory ants (Solenopsis invicta); i: predatory spiders (Nephila clavipes); j: commensal and mutualistic dipterans (Chrysomya albiceps); k: decomposing tetrapod animals (Sus scrofa domestica). Credits: a: Judy Gallagher (Wikimedia Commons) CC BY 2.0; b: pixy.org CCO; c: UBcontributor (Wikimedia Commons) CC BY-SA 4.0; d: Fedaro (Wikimedia Commons) CC BY-SA 4.0; e: Wikimedia Commons Public Domain Marck; f: United States Department of Agriculture (Wikimedia Commons) Public Domain Marck; g:Udo Schmidt (Flickr) CC BY-SA 2.0; h: Stephen Ausmus (Wikimedia Commons) Public Domain Marck; i: Ismael Etchevers; j: Hectonichus (Wikimedia Commons) CC BY-SA 4.0; k: Hbreton19 (Wikimedia Commons) CC BY-SA 3.0. All images were cropped
The eradication of a species implies a risk of secondary extinctions and cascading effects which magnitude depends on the ecological role of the species that is lost and the capacity of the rest of the species with which it interacts to compensate for its loss10)(11)(12. The potential effects of NWS loss, its ecological redundancy with other species, its effect as an indirect regulatory agent of other species, and its participation in critical ecosystem functions are analyzed below. In Figure 1, we present a conceptual synthesis of the network of ecological interactions of NWS described earlier in this work.
The NWS coexists with other species with similar ecological roles as prey and pollinator, presenting a high functional redundancy in the communities with which it interacts10)(11. This determines a high capacity of ecological communities to compensate for its loss. The strength of the interactions that provide redundancy determines the stability of communities in the face of the disappearance of a species68. The NWS is one of the calliphorid species with the lowest densities throughout its range62)(63)(64)(65. Other calliphorid species that cohabit with the NWS have similar roles as pollinators, prey, and hosts for parasitoids and phoronts, and exhibit stronger ecological interactions with partners in these interactions due to their higher densities. This set of characteristics determines a high functional redundancy in the interactions of the NWS.
Although the loss of carnivorous parasites can lead to secondary extinctions of other parasites or carnivores69, the effect of NWS as an indirect regulatory agent of other species through its role as a parasite is neutral. When a parasite exploits and regulates the populations of a particular host type, it can lead to the excessive proliferation of its host relative to other species with which it competes69. The increase in the populations of its host causes the competitive exclusion of species in the latter's trophic level, leading to they becoming rare69. Consequently, the populations of other secondary consumers, parasites or carnivores, which exclusively exploit the species affected by competitive exclusion due to the lack of prey, are also reduced69. However, for this process to occur and cause secondary extinctions, the initially lost parasite must exploit more different resources than the other species of secondary consumers. NWS parasitizes mainly medium and large herbivorous, carnivorous and omnivorous mammals, and to a lesser extent small mammals and birds, but does not show preferences for any particular host. The species competing with the main NWS hosts are other NWS hosts. Therefore, the effect of this parasite as a regulatory agent of competitive exclusion in its lower trophic levels is neutral, and its loss could not cause secondary extinctions derived from this phenomenon.
The MB does not have essential participation in critical functions of the ecosystems. Brody and others11 identified a group of critical functions for the prevention of secondary extinctions and the maintenance of the structure, biogeochemical processes and resilience of ecosystems. Such critical functions are seed dispersal, predation, disease buffering, phosphorus transport, pollination, engineer species, and foundation species11. The NWS has a role in the critical function of pollination; however, its contribution is limited or insignificant compared to that of other much more abundant calliphorids in its range that perform the same function. No other critical function is recognized in which the NWS could play an important role.
4. Conclusions and future insights
It is estimated that the NWS presents a high functional redundancy in most of its ecological interactions. Furthermore, it does not play an important role as a regulatory agent of other species and neither in critical ecosystem functions. Therefore, its eradication is unlikely to cause secondary extinctions or cascading effects in the networks of ecological interactions it integrates.
The accuracy of the potential impact assessment of the NWS eradication is limited by the information gaps that certainly exist on its ecological interactions network in Uruguay. In addition, there is a lack of monitoring data or specific research on the effects of NWS eradication in countries where such programs have already been implemented. Therefore, to opportunely detect possible impacts during the course of the program, it is recommended to monitor the ecosystems with a high spatial and temporal resolution, using indicators related to direct and indirect partners in the ecological interactions of the NWS.

