











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references

 Permalink
Permalink
1. Introduction
Livestock systems based exclusively on natural grasslands face a low forage supply during winter. This limits the growth of calves during their first winter, forcing farmers to sell them once weaned, in autumn. Lot feeding in winter and returning to grazing in the spring has been presented in Uruguay and the region1)(2 as a technological alternative that would be beneficial in areas with no agroecological aptitude for the implantation of pastures or annual crops. This would allow to exploit the growth potential of the calf and decide on the best moment for sale.
The physically effective fiber (pendf) supply in total mixed rations used in feedlots is essential for good animal performance3)(4. The pendf represents the ability to promote mastication and rumination of dietary fiber, contributing, through greater saliva production and buffer supply, to pH control and proper functioning of the rumen5. The total pendf supply from the diet depends on the total fiber content (ndf) and its effectiveness to promote rumination and mastication. This characteristic has been associated with the fiber nature and the particle size6, and it is maximum in the fiber provided by hay3. In recent years, alternative fiber options have been evaluated, since having forage reserves (hay or silage) as a source of fiber to incorporate into feedlot rations can represent a difficulty for breeder livestock systems. Studies evaluating the replacement of hay with the use of by-products that have low nutritional value but are rich in fiber, such as rice hulls and wood chips from the cellulose industry, showed the viability of removing long fiber from hay without affecting animal performance, providing enough pendf even when included in a low proportion in the ration (6% to 8%, dry matter basis)7)(8)(9.
Since whole oats (wo) are covered with hulls, they present higher fiber content (32%) than other cereals, such as sorghum (12%) or corn (10%), as well as lower starch content3. This means that the wo can be considered as another possible source of fiber in fully mixed rations for beef; however, it is unclear whether replacing the long fiber of the forage with wo could adversely affect the supply of pendf in the ration. On the other hand, unlike forage or low-quality by-products used as alternative sources of fiber, wo provide starch, which is highly degradable compared to other grains10. In this sense, the level of wo that optimizes the pendf supply may not be compatible with the increase in diet fermentability, affecting feed intake, animal growth and conversion efficiency. There is little research evaluating wo as a fiber source in calves diet11)(12; consequently, it is relevant to quantify the animal response to this management in order to generate technical coefficients that contribute to decision making, and to deepen the nutritional assessment of wo when used as fiber source in total mixed rations. The hypothesis proposed that there would be an optimal level of wo inclusion that optimizes animal production and feed use efficiency, as a result of changes in intake and associative effects between forage and grain at ruminal level that would modify the digestive use of the diet.
Two experiments were designed to characterize wo value as a source of fiber in concentrated diets for growing cattle. They aimed to assess the effect that the inclusion of wo has on growth and feed use efficiency, when replacing long fiber provided by hay (Experiment 1), and to characterize the ruminal fermentation pattern of the feed when long forage fiber is entirely replaced by wo (Experiment 2).
2. Material and methods
The experimental protocol was carried out according to recommendations of the Honorary Commission for Animal Experimentation of the University of the Republic. Experiment 2 has been approved by the Animal Ethics Committee of the Agronomy College (Res Nº 1893 CFA).
2.1 Experiment 1
Twenty-four Hereford calves (177± 18 kg) from the experimental herd of the M. A. Cassinoni Station of the Agronomy College (Paysandu) were drawn to eight groups, and these were assigned, in a random plot design, to one of the four experimental isonitrogenated rations, characterized by increasing levels of wo inclusion (variety Byzantina) replacing chopped grass hay (gh, Setaria italica) as dietary fiber sources: 0:21, 7:14, 14:7 or 21:0% (dry matter basis). Table 1 presents the ingredient and chemical composition of the four experimental rations.
Animals were held in semi-covered pens (5 m2 per calf), each containing a feeder (feeding space of 60 cm width per calf) and a drinker with ad libitum water supply. The experimental diets were gradually introduced over four weeks. During the experimental period (10 weeks), feed was offered ad libitum in three daily meals of equal amount (8 am, 12 pm and 6 pm), adjusting the supply through bunk reading to obtain 10% residual feed. Bunk reading was performed early in the morning prior to the first feeding of the day. Residual feed was discarded once weighed.
Animal live weight (lw) was recorded at the beginning of the experimental period and every 14 days, without fasting, before the first meal of the day. Rump height was also measured, both at the beginning and end of the experimental period14. Dry matter intake (dmi) was estimated daily in each pen as the difference between the offered and leftover feed (on a dry basis). Feed to grain ratio (fg) was calculated as the quotient between the daily average dmi and the daily average lw gain in each lot (wg, estimated from the lw regression on the experimental days). Duplicate samples of the offered and leftover feed were taken weekly. One of the samples was dried in a forced-air stove at 60 °C until constant weight to determine dm content; and then preserved for further chemical analysis. The second sample was used to characterize the pendf supply. The latter was quantified based on the particle size distribution in the ration using the Penn State particle separator15, and calculated as the product between the physical effectiveness factor (proportion of particles > 1.18 mm) and the ndf concentration in the whole sample6.
In week nine of the experimental period, the apparent dry matter digestibility (dmd) was estimated in vivo using the concentration of acid-insoluble ash as internal marker16. Fecal samples were collected from each animal twice a day for three consecutive days, and were combined at the end of the day in a composite sample (approximately 300 g) and frozen at -18 °C for their conservation until further processing. Samples of the offered and leftover feed were also taken for three days, starting the day before the feces sampling.
The daily intake of digestible dm was calculated as the product between the dmi and the dmd coefficient. The consumption of pendf, crude protein (cp), ndf and ether extract was calculated from the nutrient concentration (nc) in the offered feed (of) and the leftover (C, g/d = of kg/d × nc g/kg - leftover kg/d × nc g/kg). Selection in the feeder against fiber was evaluated from the difference in ndf concentration between the tmr and the leftovers per pen.
Chemical analysis. Weekly samples of offered and leftover feed were ground to 1 mm and combined into one composite sample per pen (each week contributed the same amount). Thawed fecal samples were dried to constant weight in a forced-air stove at 60 °C. Dry feces and feed samples taken during week nine for dmd estimation were ground to 1 mm and then combined into a composite sample per pen (each day contributed the same amount). The content of dm (method 934.0117), organic matter (om, method 942.0517), cp (N×6.25; method 984.1317), ether extract (method 920.3917), acid detergent insoluble N18, ndf using α-amylase and correcting by ash contamination (andfom) and acid detergent fiber (adf)18 were determined on feed samples. On the other hand, dm, om and acid insoluble ash19 were determined on feed and feces samples taken for the estimation of dmd.
Table 1: Ingredient and chemical composition of the experimental rations deferring in the level of whole oats (wo) as a fiber source replacing grass hay
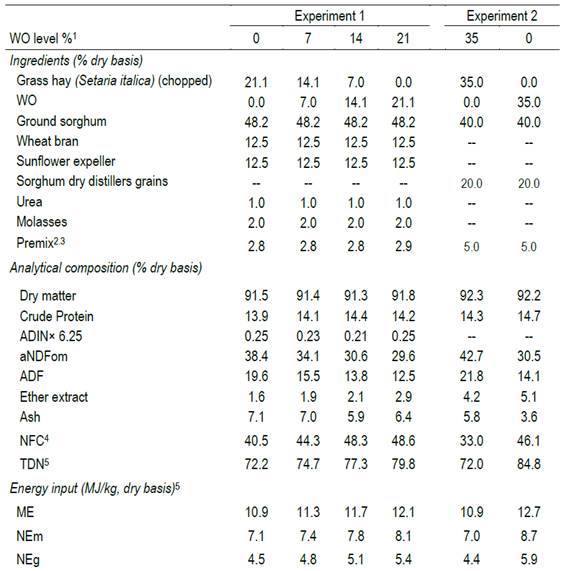
ADIN: Acid detergent insoluble Nitrogen; ADF: acid detergent fiber; aNDFmo: neutral fiber detergent using α-amylase, corrected by ash contamination. 1Whole oats replacing grass hay (dry basis); 2Experiment 1. Premix composition: calcium carbonate 61%, Zoodry feedlot (mineral - vitamins) 7%, NaCl 2.5%, Rumensin (10% monensin), 4%, Beef Sac yeast 3% (dry basis); 3Experiment 2. Premix composition: urea 19.5%, calcium carbonate 22.5%, mineral and vitamin supplement 2.3%, Rumensin (10% monensin) 0.5%, wheat bran 55.3% (dry basis); 4NFC (non-fiber carbohydrates) = (100 - NDF - crude protein - ether extract - ash)6; 5estimated values based on Beef cattle nutrient requirements model13
Statistical analysis. The experiment was analyzed according to a random plot design, taking as an experimental unit each pen with three calves, and according to the following general model Yij = β0 + τi+ εij; being: τi the effect of the i-th treatment, εij the experimental error. The lw, dmi, and pendf records were analyzed as repeated measures over time using the mixed procedure of sas (sas Inst. Inc., Cary, nc). The autocorrelation between repeated measures was modeled according to a first-order autoregressive scheme. The wg was analyzed according to a mixed linear model of slope heterogeneity of lw as a function of time, according to the general model: LWijk = β0 + Ti + εij + β1Dayijk + (β1i-β1)TiDayijk + εijk; being: Ti the effect of i-th treatment, εij the experimental error (between animals), εijk the error of repeated measurement (within animals) and slopes (β1i) representing the wg. In order to study the dmi variability, the statistical model included, in addition to the effect of the experimental week (S) and the day within week (D), the respective interactions with Ti: Yijklm = μ + Ti + εij + Sk + Dl + (TS)ik + (TD)jl + (TDS)ikm + εijklm. The fc, dmd, final lw and nutrient intake were analyzed using the glm procedure of sas (sas Inst. Inc., Cary, nc). Linear and quadratic contrasts were used to compare means associated to the wo level in the diet. The probability of occurrence of type I error was considered to have a significant effect when it was less than 5%, and to be a trend when it was less than 10%.
2.2 Experiment 2
Four Hereford steers (530 ± 100.6 kg) fitted with a ruminal cannula (KEHL®) were randomly assigned, in a crossover design, to two isonitrogenated experimental diets differing in the fiber source used: chopped grass hay (ration GH) or whole oat grain variety Byzantina (ration wo) included at a rate of 35% of the ration (dry basis, Table 1). Animals were housed in indoor individual pens with concrete floor, each containing a feeder and a drinker with ad libitum water supply, and were gradually introduced to the intake of the experimental ration. The offered feed was adjusted to 2.5% of lw, and supplied in three meals (8 am, 12 pm and 5 pm). The experimental period consisted of two periods of 21 days each, corresponding the first 14 days to the transition phase between diets, and the remaining days to measurements. During period II, animals assigned to the ration gh in period I were given the ration wo and vice versa. Animals were examined daily in order to detect any digestive or health problems.
The dmi was measured as described in Experiment 1 (day 15 to day 21), and daily samples of offered feed and leftovers were taken, which were dried in forced air stove at 60 °C for 48 h for dm content determination. Dried samples were ground to 1 mm and combined in a composite sample by animal and period. The kinetic of ruminal degradation (deg) of the dm of the ration was studied using the in situ incubation method20. Dried and ground samples (1 mm) of the feed offered in each treatment (15 mg/cm2 bag) were placed in polyester bags (9 × 11 cm, pore average size of 40 μm). These were incubated in the rumen's ventral sac before the first meal of the day (day 19), and removed in pairs (two bags per time) at 2, 4, 8, 12, 16, 24, 48, and 72 h after incubation. Bags representing t=0 for the estimation of the dm soluble fraction were only submerged in water at 39 °C for 15 minutes. Once removed from the rumen, the bags were placed in iced water to stop fermentation, and then frozen at -18 °C for preservation. After sampling, thawed bags were washed and processed according to the procedure described by Trujillo and others21, and dry weights of residues were registered. The values of deg at time “t” by animal and period were adjusted to the equation deg= a + b (1-e- kd*t)22, from which were estimated the soluble (a) and slow degradable (b) fractions, and the fractional degradation rate of fraction b (kd). Effective degradability (ed) of dm was considered as (ed= a + (b × kd)/(kd +kp)) for an estimated passage rate of kp= 0.04/h based on afrc23. Starting on day 20, ruminal fluid pH was measured every 3 h, during 24 h. Ruminal fluid samples were filtered through a double cheesecloth and pH was immediately measured using a portable pH-meter (pHTesters 20®, Oakton).
Chemical analysis. The content of dm, cp, ether extract, ndf and adf in offered feed and leftover samples was determined following the described analytical procedures in experiment 1.
Statistical analysis. The experiment was analyzed according to a crossover design in agreement with the general model Yijk = µ + Pi + Aj + Tk + Ɛ ijk, including the animal (A) as a random effect, the period (P) and treatment (T) as fixed effects, and the experimental error (ε). For pH values analysis, the fixed effect of the repeated measurement over time (sampling time) and its interaction with the treatment were also considered. The mixed procedure of sas was used, comparing the means using the Tukey test. A significant effect was considered when the probability of occurrence of type I error was less than 5%.
3. Results
3.1 Experiment 1
3.1.1 Effective fiber supply, intake and digestibility
Replacing gh with wo modified particle size distribution, linearly decreasing particles larger than 19 mm and linearly increasing particles ratio between 8.0 and 1.18 mm (P<0.01), resulting in a quadratic reduction of pendf supply (Table 2).
Table 2: Effect of replacing grass hay with whole oat (wo) as fiber source on particle size, physically effective fiber supply, intake and apparent dry matter digestibility
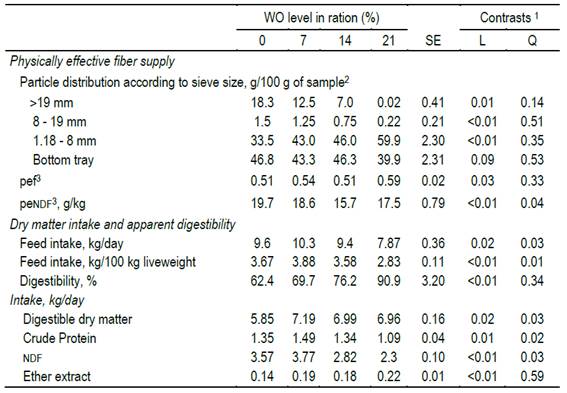
1se: mean standard error; L, Q: linear and quadratic effects of wo level on ration; 2percentage of total sample weight retained in different size meshes in Penn State Particle Separator15; 3ndf: neutral detergent fiber; pendf: physically effective fiber; pef= physical effectiveness factor (proportion of particles retained on 3 sieves, >1.18 mm); pendf g/kg=ndf, g/kg × pef6
The dmi also responded quadratically to the level of wo in the ration (Table 2), expressed both on a daily basis (dmi, kg/d= 9.65 + 0.16x - 0.012x2, R2x 0.98), and according to lw (dmi, kg/100 kg LW= 3.6 + 0.063x - 0.05x2, R2= 0.99). Likewise, a significant interaction (T×S) was recorded showing a dmi (kg/100 kg LW) more variable between weeks in wo= 21% (cv 21.3%) than in the remaining treatments (cv 6.5%). This variability in wo= 21% was given by a lower intake in the first two weeks, showing no differences in the rest of the experimental period. The (T×S) interaction for dmi (kg/d) shows cv of 11.2%, 12.4%, 12.0% and 13.0% for wo 0, 7, 14 and 21%, respectively. However, the variation between consecutive weeks was smaller in wo 21% (Figure 1). The day-within-week effect was significant (P<0.01), but independent of the treatment (P= 0.82).
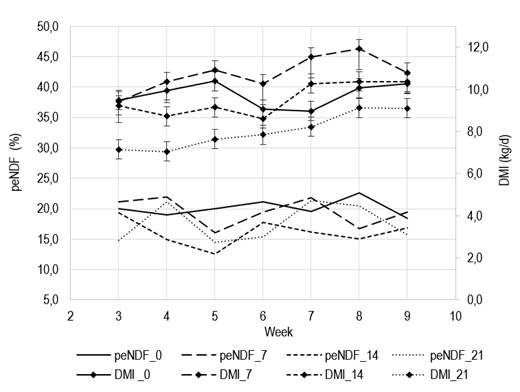
Figure 1: Effect of whole oat level (0, 7, 14 or 21%) as fiber source replacing grass hay on physically effective fiber intake (pendf), and weekly evolution of mean daily dry matter intake (dmi)
A positive linear response was observed between the dmd and the concentration of wo (dmd, %= 60.95 + 1.32x, R²= 0.96; Table 2) resulting in a quadratic increase of digestible dmi (ddmi, kg/d=0.007x2 + 0.19x + 5.94, R² x 0.87). Intakes of ndf and cp decreased quadratically (ndfi, kg/d=-0.004x2 + 0.009x + 3.65, R²= 0.91), while ether extract intake increased linearly (Table 2). The replacement of gh with wo did not affect the selection in the feeder, resulting in similar concentration of ndf between leftovers and offered feed (difference (ndf)ration-leftovers-0.4, 0.0, 0.1 and 0.4 percentage units, for WO at 0%, 7%, 14% and 21%, respectively).
3.1.2 Growth and conversion efficiency
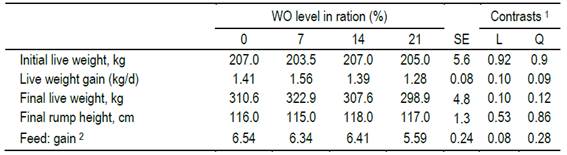
The wg was not significantly affected by the treatment (P=0.07), but showed a quadratic trend (P=0.09) increase (wg, kg/d= 1.43 + 0.02x - 0.0013x2, R2= 0.81), registering a maximum wg of 1.51 kg/d for a concentration of wo= 7.7%, from which the wg decreased, reaching for wo= 21% lower values than the control WO=0% (Table 3). However, no differences were detected in the final lw, nor in the rump height (P>0.10; Table 3). Feed conversion showed a trend (P = 0.08) to fall linearly (fc= 6.637 - 0.0397x, R2= 0.70).
3.2 Experiment 2
Replacing gh with wo did not affect the pendf supply (21.8% vs 20.4%, se 2.70, P>0.10), and no differences were observed in the physical effectiveness factor of wo compared to gh (100% vs 95%, se 1.93, P >0.10). In both treatments, no leftovers were found when feed was supplied at a rate of 2.5% of lw, finding no differences in dmi (gh: 13.7 vs wo: 13.4 kg/d, se 0.36, P>0.10). Daily average rumen pH was lower in the ration with wo (5.9 vs 6.3, se 0.036, P<0,01), staying below the pH values of the gh ration, for the 24 h. A significant effect of sampling hours was recorded (P<0.01), independent from the treatment effect (P>0.10; Figure 2). Both curves showed a similar evolution during the day; wo curve varying between 6.1 and 5.8, and the gh one between 6.6 to 6.1. Replacing gh with wo did not affect the soluble fraction of dm of the respective rations (gh: 22.6% vs wo: 20.4%, se 5.9, P=0.80), nor the potentially degradable fraction of dm (gh: 74.0% vs wo: 71.2%, ee 1.73, P=0.34). However, kd was greater for wo (0.35 vs 0.09 /h; P= 0.02), so that, for a passage rate of 0.04/h, the ed was greater for the wo ration (65.4% vs 58.3%, se 1.53, P= 0.048).
4. Discussion
4.1 Effective fiber supply and digestibility
The complete replacement of gh with wo represented a reduction of 23% of the total ndf supply in the diet. Its effectiveness to stimulate mastication and rumination was also reduced, registering a drop of 4% in the pendf concentration in the ration. However, the variation range observed when increasing the wo content in the ration (between 16% and 19% of pendf, dry basis) was above 15%, which Mertens3 suggested as the adequate value to maximize wg in finishing diets, but below 20%, indicated as adequate to reduce liver abscesses, or maximize fiber digestion4. However, substituting grass hay for wo in Experiment 2, representing 35% of the diet, resulted in optimal levels of 20% pefdn.
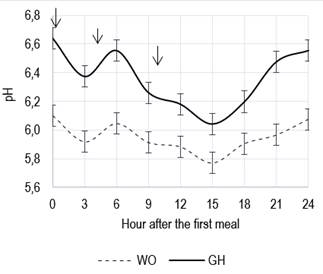
Figure 2: Daily evolution of ruminal pH in steers consuming a ration with grass hay (gh, 35% dry basis) or whole oats (wo, 35% dry basis) as fiber source (bars indicate standard error, and arrows feeding schedules)
The reduction of ndf supply by increasing wo was offset by an increase in the content of non-fiber carbohydrates and ether extract in the ration, which would partly explain the linear increase observed in the coefficient of apparent dmd in calves. When increasing wo supply, the starch supply also improved. Although starch content in this grain is low compared to other winter and summer grains (58.1%, cv 7.1%)10, it is characterized by its high digestibility (98.3%24). The improvement in dmd when replacing gh by wo was also consistent with the observed 36% reduction in the adf content of the ration wo=21%, as well as the results of in situ degradability of the dm obtained in Experiment 2. However, the magnitude of the dmd change was greater than the change in tdn concentration predicted from the model bcnrm13 (Table 1).
Although no differences were observed between rations in the potential dm degradability, the ration with wo presented a higher fermentation rate. This response could be due to not only the total removal of forage in this ration, but also to the high ruminal degradability of the dm of the wo10)(25. Studies in situ evaluating the fermentation kinetics of this grain report that 80% of the dm disappears in the first two hours of incubation in the rumen, basically associated with the high degradability of starch (96.6%); while the slowest degradable fraction would correspond to the fermentation of the grain fiber, comparatively less digestible10.
As a consequence of the higher fermentation rate of dm in the ration with wo, the effective ruminal degradability (calculated in both treatments for the same passage rate of 4%/h, given the absence of differences in dmi) was 12.2% higher compared to the ration gh. This positive effect of substituting gh by wo on effective degradability could have been accentuated in Experiment 1, where the fall in dmi when increasing wo supply above 7% of the ration (dry basis) would have contributed to a reduction in the feed passage rate, consequently increasing its retention time in the rumen. Estimates based on afrc23 show that the retention time would have increased 20.1% in wo=21% compared to wo=7%. This is relevant if the grinding of the ration samples with wo for their incubation in situ is considered to favor the expression of a high fermentation rate, probably higher than with whole grain supply, just as it was offered in the diet. In whole grain, the pericarp and hull act as a barrier for the fermentative action of bacteria on starch26, reporting a 35% dmd for the oat hulls27. Although few studies evaluate the response of whole oats to processing28, it has been reported that, in young animals that masticate sufficiently and with low levels of forage in the diet, processing is not necessary29. Morgan and Campling12 report 83% starch digestibility for whole oats offered to young cattle (7-16 months of age). The observed response of dmd in the calves of Experiment 1 agrees with this report. It is also probable that the increase in the retention time of the feed in the rumen estimated for wo=21% has favored greater mastication and rumination of the grain, facilitating the exposure of starch to the rumen microbiota30, thus contributing to a greater ruminal fermentation and the greater total dmd in this treatment.
On the contrary, at low wo inclusion levels, up to 7%, the increase in dmi and in the passage rate with respect to wo=0% may have favored a change in the site of digestion of components with a lower rate of fermentation, such as sorghum grain, moving part of the digestion to the small intestine31. This modification of the digestion site would result in an improvement in the overall energy efficiency of the growing cattle32 due to more efficient starch digestion, which could partly explain the observed trend of a higher wg of wo=7% for a similar digestible dmi in relation to higher levels of wo in the ration.
4.2 Intake and feed conversion efficiency
Although dmi presented a quadratic response with a maximum in wo= 6.9% of the ration (dry basis), then decreasing sharply until wo=21%, the concomitant linear increase in the dmd determined that the digestible dmi increased until wo=13.7%, remaining relatively constant until wo=21%. Changes in the energy concentration of the diet and its fermentative characteristics, forage relative weight and filling capacity would explain the changes observed in the dmi as the wo content increased in the ration33. On the other hand, the positive response observed in the digestible dmi up to 13.7% of wo in the ration and its subsequent stabilization would show an animal strategy tending to consume up to a constant energy level34. The strong reduction in pH values and the increase in feed fermentation rate detected in Experiment 2 when gh was replaced by wo were consistent with the responses in dmi and digestible dmi observed in the calves of Experiment 1.
Although the rations were isonitrogenous, the response in dmi meant a reduction in cp intake by increasing the wo in the ration. However, the metabolizable protein supply -estimated based on the prediction equations reported by afrc23- would have resulted in a positive balance regarding the requirement of the nutrient in all treatments, not limiting the expression of the wg potential. The wg tended to vary quadratically, maximizing (1.55 kg/d) at wo=7.7%, falling 3.4% and 15.0% when the wo content increased to 14% and 21%, respectively. Nevertheless, a proportionally larger reduction in dmi when the wo level exceeded 7.7% (6.0% and 23.1% for wo= 14% and 21%, respectively) resulted in a linear improvement in the fc as the gh of the ration was removed (Table 3). Completely replacing gh by wo improved the fc in 14.5% compared to wo 0%, evidencing an increase in the net energy for gain (neg) supply of the diet35. Even when the digestible dmi stabilized from 13-14% of wo in the ration, the improvement tendency observed in fc is consistent with the greater metabolizable energy supply (me) and neg (Table 1). With wo increase in the ration, a more efficient fermentation pattern would be expected, on the one hand, characterized by a higher propionate proportion compared to acetate and lower production of methane31)(36, and on the other, greater efficiency in the use of me for tissue deposition, due to an improvement in the quality of the diet23.
Experiment 2 results show that the substitution of gh for wo would not have predisposed the occurrence of subclinical acidosis, which also explains the improvement observed in terms of fc of the calves when replacing gh by wo. The change in the fiber source modified the fermentative characteristics of the diet, contributing to a lower average ruminal pH that would show a greater production of organic acids31. Although the daily pH curve in wo was always below that of gh, the minimum value observed (5.9) was higher than the critical value (5.6) below which the animals could experience subclinical acidosis37. A drop in the dmi associated with the increase in its variability between days could also be an indicator of possible subclinical acidosis38. On the contrary, Experiment 2 showed no differences in dmi, and there was no significant TxD interaction for dmi of the calves in Experiment 1 that would allow suspecting the occurrence of acidosis. This response suggests that the wo supply of pendf would have been adequate.
Contrary to the results obtained in the present study, other authors11 who evaluated the total substitution of alfalfa hay by wo included at a rate of 15% of the dm in a diet based on whole corn grain plus sunflower expeller offered to calves and steers found no effect on live weight gain or conversion efficiency in both categories. However, in that study, the use of whole corn grain as the main component of the ration, due to its size characteristics, would play a role in providing effective fiber to the diet, promoting insaltivation and rumen pH control39, diluting the effect of the whole oats per se as a source of pendf. In the present study, wo has shown its value as an effective fiber source in a diet without whole corn grain. Nevertheless, it should be noted that other nutritional and management aspects contemplated in the experimental approach would have contributed positively to the observed productive response. On the one hand, the basal composition of ingredients of the common ration in all treatments is highlighted, including feeds with low fermentation rate, mainly ground sorghum grain, which represents 48% of the ration and is characterized by low starch degradability10, and the use of fiber-rich by-products (sunflower expeller 40.0%, wheat bran 42.8%)40. On the other hand, to complement these, preventive measures aimed at controlling acidosis were implemented, such as the use of ionophores, ad libitum offer divided into several meals, and bunk reading41)(42, among others. These considerations are proposed by different authors as variables to consider when defining the minimum necessary pendf supply in a ration3.
5. Implications
The availability and handling of bulky feed in calf feedlot systems are often operational constraints that disable their implementation in cow-calf systems. Partial and total replacement of grass hay (up to 21% of the diet, dry basis) with wo as a fiber source in a ration based on sorghum and fibrous by-products, such as sunflower expeller and wheat bran or ddgs, has proven viable, supplying enough pendf and also energetically concentrating the diet. Given the response observed in wg and fc, the decision as to the best wo inclusion level will depend on the input: product price ratios.

