











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references

 Permalink
Permalink
Introducción
En los últimos años los cultivos energéticos se han desarrollado a nivel mundial como una alternativa viable a incorporar en la matriz energética, reemplazando combustibles fósiles y por lo tanto disminuyendo el CO2 atmosférico. Se los utiliza como fuentes de energía a través de la combustión directa, producción de gas, etanol u otro tipo de energía envasada. Otra posible ventaja atribuible a los cultivos lignocelulósicos sería la acumulación de materia orgánica en el suelo. La alta producción de biomasa aumentaría el COS, el ciclo perenne disminuiría las pérdidas de este COS a consecuencia de la respiración y erosión del suelo1)(2. Mclaughlin y otros3 observaron incrementos del COS asociado con altos rendimientos de Panicum virgatum y disminución de la erosión, debido al ciclo perenne de producción. Zan y otros4 sugieren que los cultivos perennes tienen el potencial de acumular mayor cantidad de COS que los cultivos anuales.
En Uruguay, dentro de una gran variedad de especies potenciales para generar biomasa, se iniciaron estudios en tres especies: Arundo donax (Caña común), Pennisetum purpureum (Pasto elefante) y Panicum virgatum (Switchgrass). Arundo donax es de las más utilizadas en Europa, P. virgatum está ampliamente difundida en Estados Unidos, mientras P. purpureum se cultiva en varias regiones. El ciclo estival de P. purpureum y P. virgatum es del tipo metabólico fotosintético C4, mientras A. donax es tipo C3 con desarrollo estival también1.
Es deseable que estos cultivos posean bajos valores de contenidos de humedad, ya que esto disminuye los costos de transporte, almacenamiento y secado previo a su utilización como materia prima. McLaughlin y otros3 cuantificaron para P. virgatum un contenido de humedad de 15 %. Smith y otros5 para A. donax, reportaron contenidos de 14 a 30 % de humedad, mientras Dragoni y otros6 observaron valores de hasta 61 % en este cultivo. En la especie P. purpureum se han reportado contenidos de humedad entre 657 y 78 %8)(9. El contenido de cenizas es otra variable al evaluar la calidad de la biomasa y dentro de las cenizas el componente más importante es el contenido de potasio, debido a sus efectos en la combustión3)(10.
En los sistemas de producción de cultivos energéticos los balances de energía deberían ser positivos. Los principales gastos de energía en la producción de cultivos lignocelulósicos son la fertilización, principalmente con N, de acuerdo con lo expuesto por Nassi o Di Nasso11, Fike12 y Heaton13. La fertilización con N y P, sin embargo, ha in crementado el rendimiento en estos cultivos. En P. virgatum, Searle y otros14, recomiendan fertilizar con no más de 56 kg ha-1 año-1 de N. Haque y otros15 observaron respuestas de 100 kg MSkg-1 N, mientras Pedroso16 y Owens17 observaron respuesta de 55 kg MSkg-1 N. El nivel óptimo de respuesta en biomasa total se obtuvo con 80 kgha-1año-1 N según Boehmel2. En el cultivo de P. purpureum, Sharma y otros18 obtuvieron el mayor beneficio económico con 80 kg ha-1 N y 20 Mg ha-1 de estiércol. La respuesta observada por estos autores fue de 55 a 117 kg MS ha-1 año-1 kg-1 N. En A. donax, Angelini19 obtuvieron rendimientos de 23 a 27 Mg MS ha-1 año-1 con 200 kg ha-1, 80 kg ha-1 y 200 kg ha-1, de N, P y K respectivamente. En el año 2009, Angelini y otros1 observaron rendimientos mayores, 28,7 Mg MS ha-1año-1, al fertilizar con 100 kg ha-1, 100 kg ha-1 y 100 kg ha-1, de N, P y K respectivamente.
La biomasa producida por Arundo donax, Pennisetum purpureum y Panicum virgatum podría estimarse por los componentes de rendimiento: peso individual del tallo (PIT) y número de tallos por superficie (N°Tm-2); según las investigaciones realizadas por Angelini1 en A. donax, Oliveira20 en P. purpureum y Boe21 en P. virgatum.
En Uruguay existe escasa información acerca de la producción de materia seca de los cultivos anteriormente mencionados. Es posible que estos cultivos tengan diferente potencial ya que provienen de ambientes diferentes y que la disponibilidad de los nutrientes, N y P, podría limitar la producción de biomasa en los tres cultivos. Es igualmente esperable que estos cultivos posean desarrollo radical profundo y de esta forma incidan en los contenidos de K, N y P. Se planteó como objetivo evaluar el potencial de estos cultivos en rendimiento de biomasa aérea, PIT y N°T m-2 como posibles indicadores para estimar el rendimiento, calidad de biomasa (% humedad, y contenido de K), la respuesta a la fertilización con N y P y la composición química del suelo luego de 6 años de iniciado el experimento.
Materiales y métodos
Ubicación del experimento
El experimento se realizó en la Estación Experimental «Dr. Mario A. Cassinoni» (EEMAC) de la Facultad de Agronomía, Udelar, ubicada en el departamento de Paysandú (32° 21´ S y 58° 02´W; 61 m de altitud) en Uruguay; sobre suelos clasificados como Brunosoles Éutricos Típicos Fr de la unidad cartográfica San Manuel22.
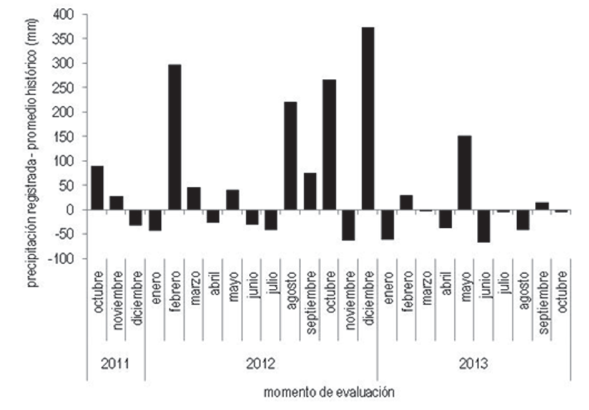
Los cultivos energéticos que se evaluaron fueron Panicum virgatum (P. virgatum), Arundo donax (A. donax) y Pennisetum purpureum (P. purpureum). Los cultivos se instalaron en primavera de 2007y se manejaron de la misma forma hasta el final del período de evaluación. El período de evaluación aquí presentado se realizó desde setiembre de 2011 a agosto de 2013. Panicum virgatum se cultivó de semilla con una población objetivo de 100 plantas m-2, P. purpureum y A. donax se cultivaron por esquejes, con una población objetivo de 2 plantas m-2. Cada parcela posee un área de 15 m2 (3 m x 5 m). Las cosechas en este experimento se realizan en agosto de 2012 y 2013. El experimento presenta un diseño en bloques completos al azar con tres repeticiones. Los datos de precipitaciones se extrajeron de Viana23.
Los tratamientos de fertilización consistieron en: 1. testigo sin fertilización; 2. 100 kg ha-1 de nitrógeno (N) agrega-do en forma de urea; 3. 100 kg ha-1 de fosfato (P2O5) agregado como supertriple; 4. 100 kg ha-1 de N más 100 kg ha-1 de P2O5. Las fertilizaciones se realizaron a un mes de realizada la cosecha (setiembre). Las fertilizaciones se realizaron anualmente desde el año 2007.
Evaluación de la producción
La cosecha se realizó con una segadora a una altura de aproximadamente 10 a 15 cm. En cada corte se cosechó un área de 2,2 m2 de cada parcela. En cada cosecha, para la determinación de materia seca se pesó una muestra fresca (aproximadamente 1 kg), se trozó a fracciones de 5 cm y secó a 60 °C durante 48 horas. El contenido de materia seca (%MS) se estimó como el cociente entre peso seco:peso fresco y se expresó en porcentaje. El contenido de humedad se estimó como la diferencia 100 - (%MS). El material secado se molió hasta pasar una malla de 2 mm y se analizó el contenido de potasio (%K). En una submuestra fresca de aproximadamente 3 kg, se pesó la muestra, se contó el número de tallos y se estimó el peso individual de los tallos (PIT), expresado en base seca. A partir del número de tallos de la submuestra se estimó el número de tallos m-2 (N°Tm-2).
Evaluación de propiedades químicas del suelo
Para evaluar en el suelo los niveles finales de K intercambiable (K int), P Bray I (P BrayI), N total (N total) y COS, se realizó el muestreo en octubre del año 2013, en el sexto año de los cultivos. Se extrajeron muestras de suelo a la profundidad de 0 a 20 cm, 20 a 40 cm y 40 a 60 cm en todos los tratamientos. Las muestras fueron secadas a 40 °C, molidas hasta pasar por malla 2 mm y enviadas al laboratorio para su análisis. La determinación de K int se realizó mediante extracción con acetato de amonio y posterior determinación en espectrometría de llama por emisión atómica24; el contenido de P Bray I se realizó de acuerdo con el extractante propuesto por Bray25 y el contenido de P en el extracto se determinó por desarrollo de color azul del complejo P-molibdato, según Murphy26; el N total se determinó por digestión húmeda de acuerdo con una modificación de la técnica de Kjeldahl propuesta por Bremner27; y COS se determinó mediante digestión húmeda según Tynsley28.
Análisis estadístico
La producción de materia fresca y seca se analizó por modelos mixtos, con el software InfoStat/P29. Al haber medidas repetidas en el tiempo se consideraron las parcelas como efecto aleatorio. Los niveles de COS, N_total, P Bray I, K_int, %K se analizaron como modelo general lineal. Se consideraron efectos significativos del ANAVA aquellos con p < 0,1 y se consideró que existió tendencia en los efectos con p < 0,2.Todas las medias se compararon por test LSD de Fisher con alfa = 0,10. La estimación de producción de materia seca a partir de PIT y N°T m-2 se evaluó por el ajuste de la regresión obtenida.
El cálculo del beneficio económico por la fertilización con nitrógeno consideró la relación: ((USD.kg-1materia seca producida)*aumento kg materia seca producida)/((USD.kg-1N invertido)*kg N invertido). Se consideró un valor de 0,1 USD.kg-1 materia seca producida y un valor de 2,17 USD.kg-1 N en la forma de urea, correspondientes a valores del año 2013.
Resultados y discusión
Producción de materia seca
La producción promedio de materia seca (Cuadro 1) fue mayor en el año 2013 (19,1 Mg ha-1) que en el año 2012 (13,7 Mg ha-1) debido, probablemente, a mejores condiciones climáticas. En el período de crecimiento 2012-2013 las lluvias se concentraron en diciembre 2012. En el período de crecimiento 2011-2012 las lluvias se concentraron en febrero 2012 (Figura 1). Esta diferencia de distribución de lluvias pudo aumentar la disponibilidad de agua durante el período de mayor sensibilidad al estrés hídrico del cultivo en el año 2013.
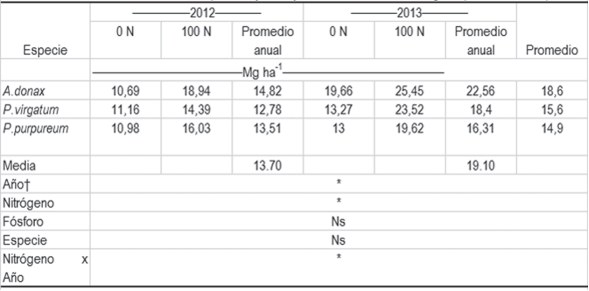
Cuadro 1: Producción de materia seca por año y manejo de fertilización con nitrógeno para las tres especies.
A pesar de que las tres especies tuvieron similares rendimientos de materia seca (p = 0,16), A. donax tendió a un mayor rendimiento promedio y P. purpureum a la menor producción (Cuadro1). Estos rendimientos estuvieron dentro de lo esperado para precipitaciones de 400 a 550 mm de acuerdo con Falasca y otros30, quienes sugieren que en Argentina A. donax puede tener rendimientos de 12 a 15 Mg ha-1. En Italia Angelini y otros1)(19 citan, para esta especie, rendimientos de 23,0 a 37,7 Mg ha-1. En P. virgatum existe en la bibliografía gran diversidad de rendimientos, según localidades y cultivares, los que van desde 5,631 a 34,6 Mg ha-1(32. Searle y otros14 sugieren que los rendimientos de este cultivo pueden ser bajos en suelos pobremente drenados y de inadecuada precipitación, y los altos rendimientos se asocian con climas cálidos, mientras los rendimientos intermedios se asocian con zonas de clima templado. Al ser nuestro clima templado a subtropical y los suelos tener un drenaje moderado a bien drenado, era esperable obtener valores intermedios de rendimiento. En la bibliografía el rendimiento de P. purpureum también es muy diverso, pero la producción de materia seca aquí obtenida se encuentra entre los registros más bajos citados. Olivo y otros33 obtuvieron, en un régimen de pastoreo, rendimientos de 23,4 a 34,3 Mg ha-1 año-1. Alencar y otros34 en cultivos bajo riego alcanzaron rendimientos de 28,2 a 35,6 Mg ha-1 año-1con fertilización de 100 a 700 kg ha-1N.
La fertilización con N aumentó, en promedio, 50 % la producción de materia seca en las tres especies (Cuadro 1), aunque al analizar por especie no hubo efecto en A. donax. En P. virgatum la fertilización con N aumentó la producción de materia seca promedio anual en 67,3 kg ha-1 kg-1 N y en P. purpureum el aumento fue de 58,3 kg ha-1 kg-1 N. Según la bibliografía era esperable que existiese respuesta a la fertilización. En P. purpureum, Sharma y otros18 reportaron respuestas del orden de 55 a 117 kg MS ha-1 año-1 kg-1 N en cultivos de P. purpureum en India, las cuales son próximas a las obtenidas en este trabajo. Las menores respuestas pudieron deberse al manejo de cosecha o pastoreo frecuente que manejaron los investigadores, acorde a lo discutido por Parrish35. En P. virgatum, Pedroso y otros16 obtuvieron una respuesta de 55 kg MS ha-1 kg-1 N en cultivos del valle central de California. Haque y otros15 obtuvieron un incremento de hasta 100 kg MS ha-1 kg-1 N en cultivos de Oklahoma. Allison y otros36 observaron incrementos de 50 % del rendimiento al fertilizar con 150 kg ha-1 N cultivos de P. virgatum en el este de Inglaterra. En A. donax el incremento de rendimiento de materia seca promedio anual fue de 69,8 kg ha-1 kg-1 N, pero esta respuesta no fue significativa. En la bibliografía se encuentran resultados dispares de la respuesta de esta especie a la fertilización. Angelini y otros19 aumentaron el rendimiento de 23 a 27 Mg ha-1 año-1 al fertilizar con 200, 80 y 200 kg ha-1 de N, P y K respectivamente, cultivos de A. donax en la zona central de Italia. Palmer y otros37 no obtuvieron respuesta o fue negativa, al fertilizar un cultivo de A. donax con 134 kg ha-1 N en la región de Carolina del Norte.
La respuesta a la fertilización con N fue mayor en el año 2013 (76 kg MS ha-1 año-1 kg-1 N) que en el año 2012 (55 kg MS ha-1 año-1 kg-1 N), debido probablemente a las mejores condiciones ambientales del 2013 comentadas anteriormente. El beneficio económico de la fertilización (asumiendo los precios a diciembre de 2013) fue de un rango entre 2,7 a 3,2. Pese a los posibles cambios en los precios de los cultivos y de los fertilizantes, el beneficio obtenido otorgaría un amplio margen de variación de precios dentro de los cuales la fertilización sería rentable.
Ninguna de las tres especies aumentó el rendimiento debido a la fertilización con P, por lo que se puede asumir que el aporte de P del suelo fue suficiente. No existe en Uruguay información que especifique el nivel crítico de P disponible para estos cultivos. La falta de respuesta a la fertilización con P pudo deberse a la adaptación de estos cultivos a ambientes de bajo aporte de Xu y otros38 sugieren que en A. donax existen microorganismos solubilizadores de P en la rizósfera, lo cual podría explicar el crecimiento de este cultivo en suelos de bajos contenidos de P disponible. Gagne-Bourgue y otros39 reportaron bacterias capaces de solubilizar P en cultivos de P. virgatum. El exudado radicular por plantas de P. purpureum podría solubilizar P ligado al Fe y Al, lo cual le permitiría adaptarse a condiciones de baja disponibilidad de P40. Al igual que en las otras especies aquí ensayadas, en P. purpureum se han identificado bacterias capaces de solubilizar P del suelo41.
El N°T m-2 y el PIT resultaron ser adecuados indicadores del rendimiento de materia seca en las tres especies. En A. donax el PIT explicó el 52 % de la variación en producción de materia seca, mientras que el N°T m-2 no tuvo una relación significativa. Al estimar el rendimiento a partir de PIT y la interacción N°T m-2*PIT, se pudo explicar el 96 % de la variación de rendimiento de materia seca. No hubo efecto de la fertilización en estos componentes. Angelini y otros1 observaron que la altura y diámetro del tallo fueron los mayores componentes del rendimiento, lo cual se puede considerar un resultado similar al aquí obtenido, ya que el PIT dependería en parte del grosor del tallo, aunque esta relación no pudo cuantificarse en el presente trabajo. En P. purpureum, a pesar de que la relación entre el rendimiento y N°T m-2 fue baja (R2 = 0,25), este fue el componente más importante del rendimiento. Al considerar un efecto cuadrático de N°T m-2 se pudo explicar el 45 % de la variación de rendimiento, por lo que el rendimiento aumentó hasta cierto valor de N°T m-2 pero luego disminuyó. Este descenso pudo deberse a que un aumento de la densidad de tallos haya provocado una mayor competencia en su crecimiento, provocando la menor producción de biomasa a igual superficie. Aunque la fertilización con N aumentó el rendimiento no afectó estos componentes, por lo que es de suponer que pudo modificar la relación hoja:tallo o la altura de los tallos al momento de la cosecha. Oliveira y otros20 determinaron una relación satisfactoria entre rendimiento y altura de los tallos, aunque no fue una variable medida en el presente trabajo, es esperable que la altura sea influenciada por N°T m-2 debido a la competencia por luz que se genera al aumentar la densidad. En P. virgatum los componentes que definen el rendimiento de materia seca son tanto el PIT (R2 = 0,44), como el N°T m-2 (R2 = 0,39), por lo que ambas variables serían de interés para aumentar el rendimiento. Al considerar ambas variables en un modelo de regresión pudo explicarse el 93 % de la variación de rendimiento. Boe y otros21 observaron resultados similares para esta especie, ya que las variables de mayor relación con el rendimiento fueron N°T m-2 y peso del fitómero. El N aumentó el rendimiento de esta especie, lo que incrementó ambos componentes de rendimiento.
Contenido de humedad
La especie que tuvo el menor contenido de humedad a la cosecha fue P. virgatum (16 %), mientras P. purpureum tuvo el mayor contenido de humedad (52 %) y A. donax un valor intermedio (32 %). Al investigar estos cultivos con fines energéticos, McLaughlin y otros3 cuantificaron para P. virgatum un contenido de humedad de 15 % y Bulpitt y otros42 para A. donax un contenido de 28 %. Dragoni y otros6 observaron valores superiores de humedad, de 61 a 48 %, al momento de cosecha de estos materiales. Los niveles de humedad reportados en la bibliografía para P. purpureum, son ligeramente superiores a los aquí observados, debido probablemente a que estos cultivos fueron manejados con cortes en estado vegetativo 7 o cada 60 días8)(9. Es deseable que estos cultivos posean bajos contenidos de humedad, ya que esto disminuye los costos de transporte, almacenamiento y del secado previo a su utilización como materia prima para energía, excepto que sea utilizada para biogás.
El contenido de humedad promedio de las tres especies fue menor en el año 2013. No hubo efecto de la fertilización con N, ni con P, en el contenido de humedad en ninguna de las tres especies. Allison y otros36, sin embargo, cuantificaron aumentos en los contenidos de fibra en detergente neutro y lignina en cultivos de P. virgatum fertilizados con N, lo que provocaría mayor contenido de materia seca. Flores y otros43 no encontraron efecto de la fertilización con nitrógeno en los contenidos de fibra de cultivos de P. purpureum.
Carbono en el suelo
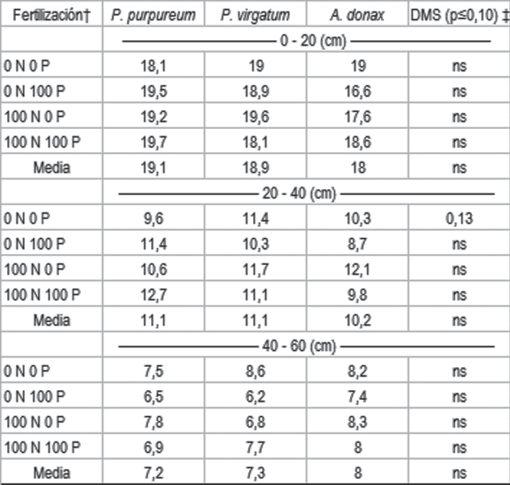
Al sexto año de los cultivos no hubo diferencias en los contenidos de carbono del suelo entre especies ni manejos de fertilización, en las profundidades evaluadas. A la profundidad de 20 a 40 cm, sin embargo, la especie P. virgatum presentó mayor contenido de COS que P. purpureum, en las parcelas sin fertilizar con N ni P (Cuadro 2). Para la profundidad de 0 a 20 cm el contenido promedio de COS, de todo el ensayo, fue de 18,6 g kg-1, en la profundidad de 20 a 40 cm el contenido fue de 10,8 g. kg-1 y para la profundidad de 40 a 60 cm fue de 7,5 g kg-1. Ji44, Lee y otros45 analizaron aumentos del carbono en el suelo al fertilizar P. virgatum durante cinco años. Jung y otros46 registraron aumentos en el COS en dos de tres sitios ensayados en Ohio, tras dos años de evaluación. El contenido de COS hasta un metro de profundidad fue mayor bajo cultivo de P. virgatum que en praderas de gramíneas perennes, pero no difirió en los primeros 15 cm47. Dou48, Lobo49 concluyen que P. virgatum puede favorecer el aumento de COS al determinar mayores valores que en cultivos convencionales. Schmer y otros50 al cabo de 10 años de cultivos de P. virgatum, en Nebraska, encontraron incrementos de 0,5 Mg año-1 de COS en los primeros 30 cm de suelo. En Texas, el cultivo de A. donax presentó, al cabo de 40 años, mayor contenido de COS hasta los 50 cm de profundidad que los cultivos de Cynodon dactylon51.
Fósforo en el suelo
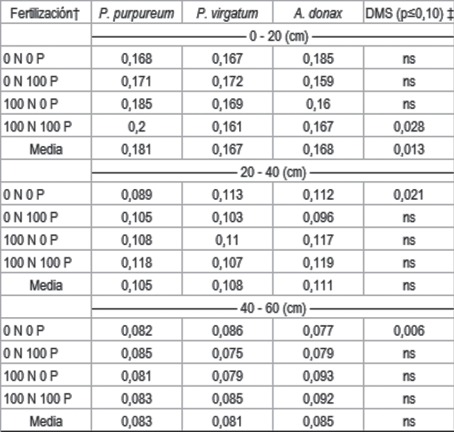
Hubo un claro efecto de aumento del P_BrayI al fertilizar con P, en la profundidad de 0 a 20 cm. Panicum virgatum fue la especie en la cual se midió mayor nivel de P_BrayI, a la profundidad de 0 a 20 cm en el tratamiento con fertilización con P. A. donax y P. purpureum presentaron similares niveles (Cuadro 3). El mayor valor analizado en el cultivo de P. virgatum pudo deberse a la menor extracción de P realizada, debido al menor rendimiento. Es de esperarse además que este cultivo haya realizado mayor reciclado del P absorbido al senescer y depositarse las hojas en la superficie del suelo. Siri y otros52 observaron menores concentraciones de P en la biomasa cosechada de la especie P. virgatum, que en las otras dos especies aquí evaluadas. Schmer50, sin embargo, observaron disminuciones de la disponibilidad de P luego de 10 años de cultivos de P. virgatum. La fertilización con N no provocó cambios en los niveles de P BrayI, pese a haber provocado diferencias de rendimiento. Es posible que la mayor cantidad de biomasa removida no provocase mayor extracción de P si la concentración de P en la biomasa fuera menor a mayor cantidad de esta.
En las otras dos profundidades de muestreo no se observaron diferencias entre especies en concentración de P en suelo, ni por causa de la fertilización con P ni N (Cuadro 3). Es posible que la diferencia observada a menor profundidad se deba a la estratificación provocada por la absorción de P por la planta y la deposición del mismo en superficie a través de la senescencia de tejido. La ausencia de laboreo, dado que son cultivos perennes, no favorecería la reincorporación de este elemento en profundidad.
Potasio en el suelo
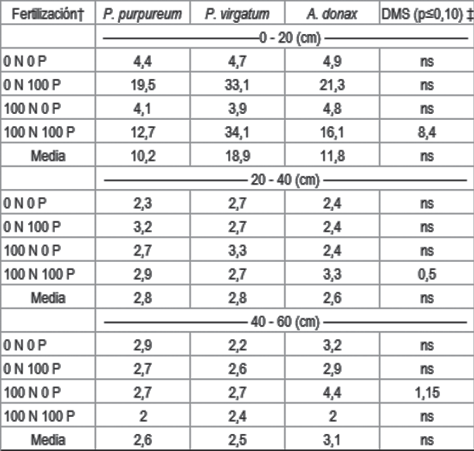
Las parcelas cultivadas con A. donax fueron las que tuvieron mayor K_int al final del período de evaluación 0,63 meq.100 g-1, de 0 a 20 cm de profundidad. Pennisetum purpureum y P. virgatum presentaron similares valores de K_int, 0,40 y 0,42 meq.100 g-1 respectivamente.
Las parcelas fertilizadas con N presentaron menores niveles de K_int, a la profundidad de 0 a 20 cm, que las parcelas sin fertilizar (Cuadro 4). No hubo efecto de la fertilización con P. Las parcelas fertilizadas con N provocaron mayor extracción de K y el K_int podría haber disminuido. Este efecto de la fertilización fue significativo en las dos especies donde se observó respuesta a la fertilización con N, P. virgatum y P. purpureum. En A. donax no hubo respuesta en rendimiento a la fertilización con N y el K_int tampoco fue diferente entre parcelas testigo o fertilizadas.
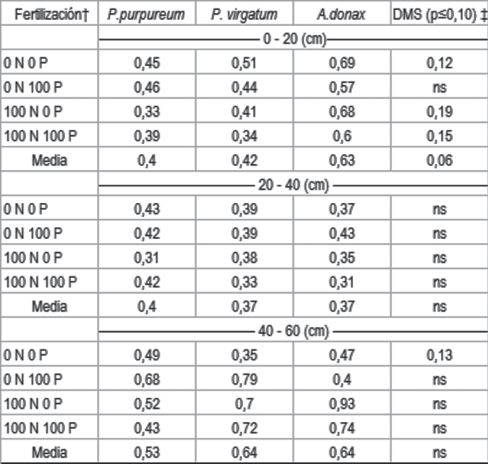
Cuadro 4: Contenido de K intercambiable (meq.100 g-1) en el suelo para cada especie, según nivel de fertilización con N y P, por profundidad de muestreo.
Al igual que en P_BrayI, en las profundidades mayores a 20 cm no se observaron diferencias en K_int entre especies ni a causa de la fertilización (Cuadro 4). Las razones de este comportamiento serían explicadas por la misma razón que lo observado en P. En los tratamientos sin fertilización, sin embargo, el suelo cultivado con P. virgatum tuvo menores contenidos de K que P. purpureum a la profundidad de 40 a 60 cm. Esto podría deberse a una mayor extracción de K por P. virgatum a esta profundidad.
Potasio en la biomasa
La especie P. purpureum fue la que tuvo la mayor concentración de K, 1,45 % (Cuadro 5). Pennisetum purpureum tuvo altos contenidos de humedad al momento de la cosecha, pudiéndose suponer que el proceso de translocación de los minerales no ocurriese de forma eficiente en esta especie. Los menores contenidos de K en P. virgatum podrían atribuirse a lo mencionado por McLaughlin y otros3, quienes observaron que en esta especie la alta relación de tallo:hoja puede favorecer los bajos valores de K en la biomasa, pero esta relación no se cuantificó en este experimento.
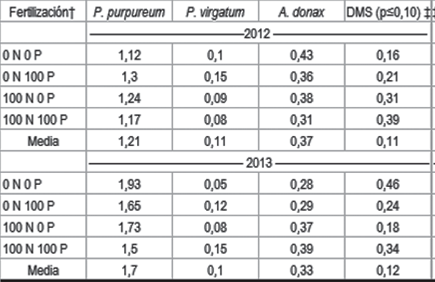
La mayor extracción de K la presentó P. purpureum, 216 kg ha-1 año-1 y la menor extracción la presentó P. virgatum, 16 kg ha-1 año-1. A pesar de la alta extracción de P. purpureum, esto no se reflejó en menores valores de K_int en el suelo, que en los cultivos de P. virgatum. Es posible que el reciclaje de biomasa senescente sea importante en P. purpureum y contribuya en mantener los contenidos de K en el suelo a la profundidad muestreada. Arundo donax extrajo 65 kg ha-1 año-1 y fue la especie que presentó mayor contenido de K_int en suelo, lo que podría atribuirse al mayor reciclado que generaría este cultivo, al depositar sobre la superficie parte del K absorbido, a través de la senescencia de las hojas. Los valores de exportación de K, en biomasa aquí observados, tienen la misma tendencia y orden de magnitud que los publicados por Siri52.
La concentración de K en la biomasa, de P. purpureum, cosechada en 2013 fue mayor a la del año 2012 (Cuadro 5). Es posible que las menores precipitaciones desde enero a marzo del año 2013 provocasen esta mayor concentración de K, al presentar mayor estrés hídrico el cultivo durante el período de translocación. Nassi o Di Nasso y otros11 observaron un incremento de K en la biomasa de A. donax debido al estrés hídrico. Este estrés también se vio reflejado en el menor contenido de humedad de los tejidos en el año 2013. Es posible que la formación de heladas haya ocurrido en la segunda semana de abril del año 2013, mientras que en el año 2012 estas ocurriesen en la cuarta semana, lo cual se presupone por las temperaturas mínimas próximas a 5 °C medidas en casilla meteorológica. Las heladas interrumpirían el proceso de translocación de K desde la parte aérea a los órganos de reserva de estas especies, por lo cual la biomasa cosechada en el 2013 tendría mayor contenido de K.
Nitrógeno en el suelo
El contenido de N_total en los primeros 20 cm de profundidad de muestreo fue mayor en el suelo cultivado con P. purpureum que en las otras dos especies (Cuadro 6). Lobo49 observó mayor acumulación de N orgánico en los primero 5 cm de suelo cultivado con P. virgatum que en suelos cultivados con especies agrícolas anuales, en tres de los cuatro sitios evaluados.
A la profundidad de 20 a 40 cm el contenido de N total fue mayor en las parcelas fertilizadas con N. Las parcelas sin fertilización con P ni N, cultivadas con la especie P. purpureum, fueron las de menor contenido. A la profundidad de 40 a 60 cm la especie A. donax sin fertilización con N ni P fue la que presentó menor contenido de N total en el suelo. Estos resultados se deberían a diferencias en el crecimiento radicular entre las especies. No se abordaron en este estudio indicadores que permitan concluir sobre este tema.
Conclusiones
Arundo donax produjo significativamente más biomasa área (18,6 Mg ha-1año-1) que P. virgatum y P. purpureum, siendo estos rendimientos obtenidos en las tres especies esperables para nuestras condiciones ambientales. En las especies A. donax y P. virgatum podría estimarse el rendimiento a partir del N° Tm-2 y peso de los mismos. Sin embargo, en P. purpureum a partir de estos componentes no se obtendría información suficiente. La fertilización con N aumentó el rendimiento significativamente en P. virgatum y P. purpureum, pero no en A. donax. No hubo respuesta a la fertilización con P, por lo que se puede considerar que el aporte del suelo fue suficiente. El aumento de P Bray I en la superficie del suelo en los tratamientos fertilizados con P se debería a que la extracción de los cultivos y la fijación del suelo fueron menores al aporte de P por fertilización. Pese a los rendimientos intermedios de P. virgatum, esta especie fue la que presentó menor contenido de humedad en la biomasa cosechada, lo que es una característica favorable como materia prima para la producción de energía. Pese a ser cultivos perennes de alta producción de biomasa, esta mayor producción no contribuyó a mayores contenidos de COS y nitrógeno total del suelo en estos primeros 6 años de estudio. Existieron diferencias significativas en la extracción de K y no hubo una clara relación con los valores de K_int, por lo que es necesario profundizar en la dinámica de su absorción y deposición en la superficie del suelo a través de la senescencia de los tejidos.

