











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references

 Permalink
Permalink
Introducción
El uso del suelo es un aspecto central para el desarrollo agropecuario y tiene influencia sobre la matriz de alternativas productivas y aspectos económicos. Las plantaciones forestales comerciales (PFC) ocupan porciones muy relevantes en la región de pastizales de la cuenca del Río de la Plata, actualmente, el área de PFC en el Uruguay supera las 990.774 ha1)(2)(3 y los impactos sobre el ecosistema campestre han sido poco estudiados.
El establecimiento de las PFC comienza con la eliminación de las comunidades nativas en las áreas destinadas a los rodales. La plantación forestal sobre comunidades nativas puede contribuir a la producción animal, si dichas comunidades son preservadas en la entrelínea4. La convivencia de áreas forestadas y no forestadas determina un gradiente de intervención desde la sustitución de la vegetación residente por especies forestales hasta la interferencia en el ambiente lumínico de las áreas no forestadas. El grado de atenuación de la Radiación Fotosintéticamente Activa (RFA) varía entre especies forestales, dependiendo de la transmisividad de sus hojas, así la cantidad de luz que llega al piso forestal dependerá del tipo de dosel que esta atraviesa, de la densidad de plantación, su altura y edad5)(6. Las prácticas silviculturales afectan la competencia, al modificar principalmente el porcentaje de cobertura de copas y otros parámetros estructurales del canopeo como el Índice de Área Foliar (IAF)6)(7)(8. El sombreado afecta la calidad, producción y composición de la pastura, en pasturas nativas se observó un incremento de las especies forrajeras valiosas asociado a la cobertura arbórea9)(10. Existen especies que reaccionan ante el sombreado disminuyendo linealmente su crecimiento, en cambio otras reaccionan ante la sombra de acuerdo a la disponibilidad de agua11)(12.
Las propiedades químicas del suelo son modificadas por la introducción de los arboles, en este sentido se han constatado una disminución del pH, aumento de la acidez intercambiable (Al+3) y disminución del contenido de las bases Ca, Mg y K de los suelos debido al cambio en el uso del suelo pasando de un sistema pastoril a forestal13)(14)(15. A simismo, los árboles también contribuyen al secuestro de carbono en el suelo16)(17, destacándose las plantaciones con destino a aserrío las cuales contribuirían en mayor medida a la mitigación de la producción de gases de efecto invernadero, aunque los volúmenes por superficie en plantaciones con destino a la producción de pulpa son mayores.
Otros de los de los efectos de las PFC son las producidas sobre el recurso agua, según Silveira18, se observó que los escurrimientos anuales en una cuenca forestada tienden a disminuir entre un 22 y 31 %, dependiendo de la precipitación anual, también aumentan la intercepción del agua de lluvia, disminuyendo los procesos erosivos pero tienden a tener más pérdidas por evapotranspiración y a modificar la dinámica del agua disponible en el suelo que una pastura bajo pastoreo19)(20)(21.
Cuando se instalan los sistemas silvopastoriles (SSP) interviniendo sobre formaciones nativas, se produce un cambio en la composición específica de la pastura, a partir de la heterogeneidad espacial que genera la intervención22. Martínez y otros23 estudiando un SSP en un semillero de Pinus taeda con densidad de 109 árboles ha-1 determinaron la dominancia de Cynodon dactylon (L) Pers. y tipos productivos ordinarios frente a los finos y tiernos. También Zarza24 identificó a C. dactylon dentro del grupo de gramíneas asociadas al efecto del monte más adaptadas a la sombra junto a Bromus catharticus Vahl. mientras que Paspalum notatum Alain ex Flüggé presentó comportamiento intermedio, Axonopus fissifolius (Raddi) Kulhm. (Axonopus affinis Chase) y Juncus sp. se ubicaron en la zona de pleno sol. Picasso y otros10 detectaron diferencias significativas del efecto de los árboles nativos sobre la composición botánica de pasturas naturales, con mayor frecuencia de especies invernales asociadas al ambiente del árbol.
Para las condiciones de Uruguay no existe información que caracterice el valor pastoral de la vegetación del sotobosque y de las áreas no plantadas en PFC para las diferentes modalidades de producción, requiriendo de esa información para sustentar el desarrollo de futuros SSP.
El objetivo del presente trabajo fue estudiar el efecto de la PFC sobre la cobertura del suelo, los tipos productivos y el valor pastoral del estrato herbáceo del rodal, callejones cortafuego y el campo nativo aledaño.
Materiales y métodos
El clima de la región es templado cálido (Subtropical húmedo) tipo Cf (Cfa) del sistema de clasificación de clima de Köppen - Geiger25.
Los sitios de estudio abarcaron tres localidades:
En cada rodal se midió densidad de árboles y sus características dasométricas (CD): diámetro a la altura del pecho (DAP), diámetro basal de copa (DBC), altura total (AT) y altura de inserción de la última rama verde (AURV), calculando con AT y AURV la longitud de copa verde (LCV) y con esta el volumen de copa27, utilizándose un medidor Vértex, una cinta diamétrica milimetrada y una cinta convencional.
La estructura de la vegetación en cada sitio se describió a través de treinta cuadros de 0,25 m2 distribuidos cada un metro sobre una transecta. Bajo el rodal los cuadros se dispusieron en el sentido de la entre fila de los árboles en las orientaciones N-S y E-O, en los callejones cortafuego y el campo natural las transectas se localizaron en las mismas orientaciones. En cada cuadro se determinó la cobertura en porcentaje28 de la cubierta vegetal (CV), suelo descubierto (SD), mantillo + heces (M+H) y restos forestales (RF).
La cubierta vegetal (CV) corresponde a la suma de las coberturas relativas de cada una de las especies, el suelo descubierto (SD) se estimó como la superficie de suelo descubierto y piedras, el área ocupada por mantillo y heces (M + H) corresponde al área no recubierta por vegetación ni suelo descubierto. El área ocupada por los restos forestales (RF) estima el área de restos forestal depositados sobre el suelo. Los relevamientos florísticos y mediciones dasométricas se realizaron entre los meses de enero y marzo del 2010.
Se analizó la calidad forrajera global de cada sitio de muestreo a través del Valor Pastoral corregido (VPc)29)(30. Esta variable se compone de: 1) cuantificación de la composición botánica, 2) asignación a cada especie de un valor relacionado con su TP31)(32 el cual varió de 0 a 10 y 3) cálculo del valor pastoral (VP) y corregido por CV (VPc).
Las equivalencias de los TP con el Índice Pastoral (IP) se presentan en el Cuadro 1.
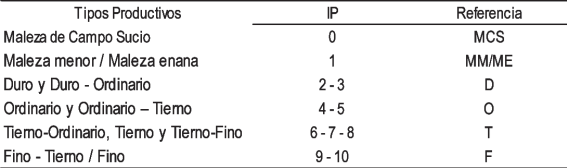
Para el cálculo del valor pastoral (VP) y el VPc se emplearon las siguientes fórmulas:
VP = Σ (% Cobertura especifica * Valor Índice Pastoral)
VPc = (VP (Comunidad) * % Cobertura Vegetal)/100
Se realizó un análisis multivariado de conglomerados y componentes principales. Los análisis de agrupamientos fueron realizados entre las 18 unidades de muestreo (sitios), considerando las CD, CV, SD, M+H, RF, TP y VPc. En todos los agrupamientos, las variables fueron estandarizadas, restando la media de cada variable y dividiéndola por el desvío estándar de la misma. La unidad de distancia entre unidades de muestreo fue la distancia euclidiana con las variables estandarizadas, y el método de agrupamiento usado fue el de Ward. En todos los casos, se probó la existencia de un número de grupos nítidos en la estructura de los agrupamientos. La prueba de nitidez de los grupos fue realizada mediante test de aleatoriedad (bootstrap), utilizando 1000 repeticiones. Para los análisis se utilizaron los software MULTIV33 e Infostat34.
Resultados y discusión
Caracterización del componente arbóreo
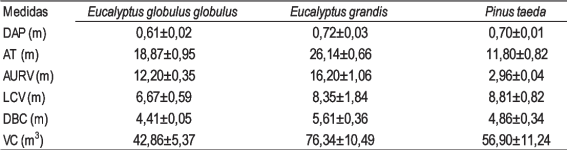
Los sitios de muestreo caracterizados por las variables dasométricas permiten discriminar tres grupos nítidos (Figura 1). Los agrupamientos fueron testados no rechazándose el número de grupos previsto siendo el p-valor: 0,398. Esas diferencias en las variables dasométricas pueden afectar las características del dosel incidiendo en la trasmisión de la radiación a través del mismo35. La distribución de la biomasa en Eucalyptus dunnii Maiden presentó un 73 % de la misma alojada en las trozas y un 27 % en los restos pos cosecha (despunte, corteza, ramas finas y hojas), en tanto E. globulus presentó un 78 % de la biomasa alojada en las trozas y un 22 % en los restos pos cosecha36)(37. La partición diferente de biomasa aérea entre especies con menor proporción destinada a la copa, podría redundar en un mayor pasaje de luz al sotobosque.
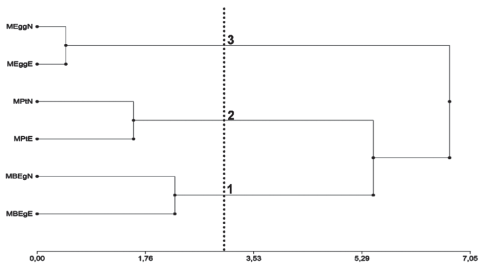
Figura 1: Dendrograma de las características dasométricas según especie forestal. Se diferencian tres grupos nítidos con un P-valor = 0,398.
En el Cuadro 2 se presentan las medias de las variables dasométricas de los grupos formados en el análisis de conglomerados. El grupo 1 está integrado por rodales de E. grandis (MBEgN y MBEgE), los cuales presentaron un mayor valor de AT y VC, el segundo grupo formado por los rodales de P. taeda (MPtN y MPtE), los cuales se caracterizaron por el menor valor de AT y AURV y el tercer grupo integrado por los rodales de E. globulus (MEggN y MEggE), que presentaron los menores DAP, LBC y VC. Los rodales de E. grandis y P. taeda presentan una mayor similitud debido a la LCV, DBC y al VC, estas características generan una mayor competencia por luz6)(7)(8)(38.
Componentes de la cobertura del suelo
En función de los sitios de muestreo y las variables área cubierta por Restos Forestales (RF), Suelo Descubierto (SD), Heces + Mantillo (H + M) y Cubierta Verde (CV) se identificaron tres grupos nítidos de sitios. Los agrupamientos fueron testados no rechazándose el número de grupos previsto siendo el p-valor: 0,43.
El primer grupo integrado por los sitios ubicados en los rodales de E. globulus (MEggN y MEggE) y callejones cortafuego (CcSN y CcSE), campos naturales de Sarandí del Quebracho (SCNN y SCNE), callejones cortafuego de E. grandis (CcBN y CcBE), campos naturales de Bañados de Medina (BCNN y BCNE) y campos naturales de Paso Bonilla (PCNN) y PCNE), con valores promedio de CV de 86,59 % (DE ± 15,80), SD de 0,15 % (DE ± 0,19), H + M de 1,23 % (DE ± 0,93) y RF de 12,03 % (DE± 15,33).
Un segundo grupo de sitios formado por los callejones cortafuego en rodales de P. taeda (CcPN y CcPE), con CV de 41,93 % (DE ± 18,28), SD de 12,29 % (DE ± 1,93), H + M de 10,25 % (DE ± 0,28) y RF de 35,53 %(DE ± 20,48).
El tercer grupo integrado por los sitios ubicados en rodales de E. grandis (MBEgN y MBEgE) y P. taeda (MPtN y MPtE), con CV de 8,27 % (DE ± 3,80), H + M de 0,88 % (DE ± 1,37) y RF de 90,85 % (DE ± 3,56), donde el SD fue cero. Este resultado concuerda con lo reportado para las variables dasométricas, donde E. grandis y P. taeda presentan menores diferencias entre ellos que las reportadas para E. globulus.
El análisis de Componentes Principales (CP) resume en dos ejes el 98,7 % de la variación total (Figura 3), agrupando los sitios según condiciones similares.
Sobre CP 1 y hacia valores negativos se ubica el segundo grupo, correspondientes a callejones cortafuego de P. taeda (CcPN y CcPE), asociadas a las variables SD y H + M. Estos callejones cortafuego tienen un ancho de 4,66 m, distancia que determina una alta incidencia del rodal sobre el ambiente lumínico, que estaría explicando la asociación a valores crecientes de SD.
El grupo integrado por los callejones cortafuego en rodales de E. grandis (CcBN y CcBE) y rodales de E. globulus (MEggN y MEggE) presentaron un comportamiento promedio para las variables (SD, H + M, RF y CV) respecto a los dos CP. Este grupo integra el grupo 3 del dendrograma (Figura 2). Las asociaciones entre rodales (E. globulus) y callejones cortafuego (E. grandis) puede ser explicado por los menores niveles de residuos forestales depositados por E. globulus debido a la relación fuste/copa37. Este comportamiento estaría explicado por los mayores niveles de RF volcados por E. grandis bajo el rodal (mayor volumen de copa, corteza caduca y mayor tasa de recambio foliar) y la deriva de estos RF hacia los cortes, independientemente de la orientación de las hileras36)(37)(39)(40.
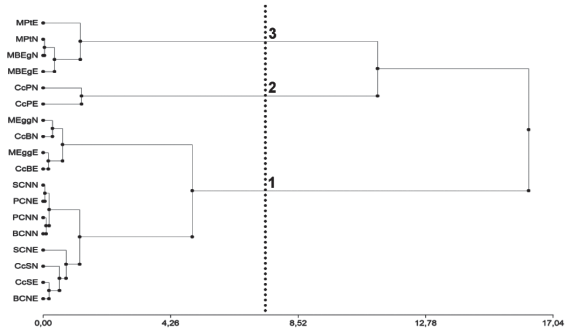
Figura 2: Dendrograma de los sitios en función del área cubierta por los distintos componentes que cubrían el suelo. Se diferencian tres grupos nítidos con un P-valor = 0,43.
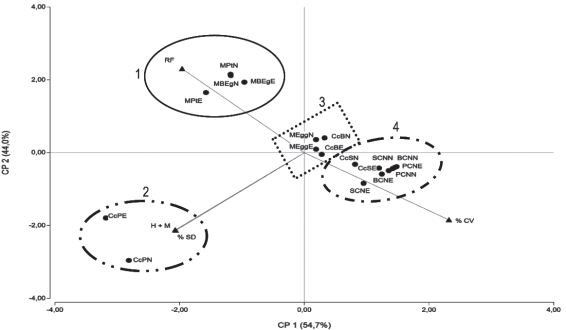
Figura 3: Biplot correspondiente al análisis de componentes principales (CP) basado en el área cubierta por los distintos componentes que cubrían el suelo (▲) según sitio (•).
En el CP 1 y hacia valores positivos se ubica un cuarto grupo caracterizado por sus mayores valores de CV, que variaron entre 85,70 y 100 %, conformado por los sitios sobre campo natural, Bañados de Medina (BCNN y BCNE), Sdí del Quebracho (SCNN y SCNE) y Paso Bonilla (PCNN y PCNE) y los callejones cortafuego de E. globulus (CcSN y CcSE). Estos sitios se asemejan en los porcentajes de CV (99,64 y 99,26 %) y H + M (0,36 y 0,65 %) para BCNN y PCNN, de CV (99,83 y 100 %) para SCNN y PCNE y de CV (98,64 y 96,94 %) y SD (0,00 y 0,06%) para BCNE y SCNE. En cambio el sitio CcSE presenta valores similares en CV (95,08 %) lo cual es explicado por el mayor ancho del callejón cortafuego (26 m) e intermedio en SD (0,03 %), a diferencia el sitio CcSN se agrupa a la mayor distancia lo cual es debido a los valores de la CV (85,70 %) y RF (12,37 %), lo que se podría explicar por la distancia existente entre rodales (11,5 m) para esa orientación. Esta menor distancia entre rodales determinaría un ambiente menos iluminado con efecto sobre la comunidad herbácea.
Sobre el CP 2 y hacia valores positivos se ubica el primer grupo que incluye los sitios situadas bajo montes de E. grandis y P. taeda, asociadas al alto recubrimiento de restos forestales. Las diferencias de esta variable entre esas especies y E. globulus se asocia a un mayor recambio foliar, desrame y descortezado. Especies forestales diferentes pueden mantener biomasas de diferente longevidad, asociados a una alta productividad y menor duración de vida de la biomasa vegetal36)(37)(39)(40.
Existen diferencias especificas en la distribución de materia seca de las estructuras que conforman el árbol, E. globulus presenta 78 % de la materia seca aérea alojada en el fuste, siendo el resto ramas menores y hojas, mientras en E. grandis se aloja en el fuste menos del 73 %, por lo que en esta especie aumenta la incidencia de los tejidos con mayor tasa de recambio, afectando los volúmenes de restos senescentes que son depositados sobre el suelo36)(37. En este mismo sentido, trabajos realizados por Muñoz y otros41 en E. nitens indican que el área foliar proyectada fue máxima con densidades de 400 árboles ha-1, mientras el índice de área foliar aumenta hasta 800 árboles ha-1 y disminuye con densidades mayores, debido a la menor proporción de biomasa localizada en la copa. Estos resultados explicarían que en rodales de E. globulus con poblaciones de 1250 árboles ha-1 existe mayor pasaje de radiación hacia el sotobosque determinando una mayor cubierta vegetal en comparación a las encontradas en E. grandis y P. taeda.
Tipos productivos
La caracterización de los sitios de muestreo por los TP de las especies permitió identificar cuatro grupos nítidos (Figura 4). Los agrupamientos fueron testados no rechazándose el número de grupos previsto siendo el p-valor: 0,099.
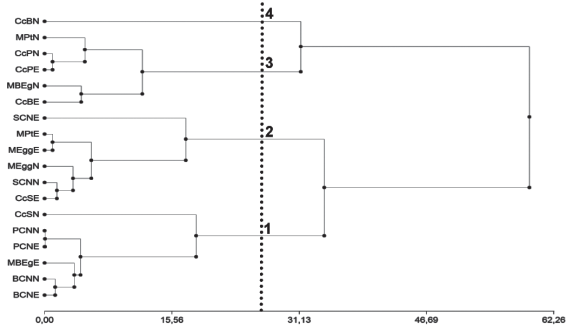
Figura 4: Dendrograma de los sitios en función de la cobertura de los TP presentes. Se diferencian cuatro grupos nítidos con un P-valor = 0,099.
En el grupo 1 los sitios PCNN y PCNE son los que se encuentran a una menor distancia, y se caracterizan por la similitud en los valores de todos los TP de las especies relevadas. Donde predominan los pastos tiernos (T), (cobertura promedio de 50,21 %), y las principales especies son: P. notatum (24,73 %), A. fissifolius (15,42 %), Axonopus argentinus Parodi (5,37 %) y Steinchisma hians (Elliott) Nash (Panicum milioides Nees ex Trin.)(3,44 %). En comunidades de campo natural la orientación de los sitios no generó diferencias en los tipos productivos ya que estaban en el mismo ambiente.
Los sitios BCNN y BCNE, presentan la mayor diferencia para el TP T (9,16 %), sin embargo esta clase es la más importante en cuanto a su contribución (54,7 % promedio). Las mínimas diferencias se dan entre los TP MCS y ME/MM (0,20 y 0,48 %) las que tienen una cobertura de 6,88 y 3,39 %, respectivamente. La principal especie MCS es Panicum milioides Malme con una cobertura de 6,47 y 6,65 %, (BCNN y BCNE, respectivamente) mientras que para el TP ME/MM se relevaron 31 especies entre las que se destacan por su cobertura Aspilia montevidense var. setosa (Griseb.) Cabrera (Aspilia setosa Griseb.) (0,77 y 0,76 %), Dichondra microcalyx (Hallier f.) Fabris (0,49 y 0,70 %), Juncus capillaceus Lam. (0,69 y 0,71 %) y Verbena litoralis Kunth (0,20 y 0,29 %). A una distancia mayor se encuentra el sitio MBEgE, con valores similares de los TP MCS y D (6,62 y 12,92 %), siendo E. horridum la única especie MCS, mientras que para el TP D fueron Nassella charruana (Arechav.) Barckworth (Stipa charruana Arechav.)(8,44 %), Paspalum quadrifarium Lam. (3,58 %) y Saccharum angustifolius (Nees) Trin. (Erianthus angustifolius Nees) (0,9 %). Este sitio presenta el valor más alto en el TP T para este grupo (65,40 %) lo cual es explicado principalmente por la presencia de Mnesithea selloana (Hack.) de Koning & Sosef (Coelorachis selloana (Hack.) A.Camus) (23,87 %), Nassella neesiana (Trin. & Rupr.) Barkworth (Stipa setigera J. Presl.) (19,94 %) y Piptochaetium stipoides (Trin. & Rupr.) Hack. ex Arechav.(17,81 %), pero esta presencia de TP T está afectada por el bajo porcentaje de CV (13,93 %). Por último, a la mayor distancia se agrupa la transecta CcSN, por similitud de los valores en los TP T y F (59,78 y 2,64 %), siendo las especies tiernas más importantes A. fissifolius (33,98 %), P. notatum (15 %) y Setaria setosa (Sw.) P. Beauv.(Setaria vaginata Spreng.) (2,77 %), mientras que la única especie fina relevada fue Paspalum dilatatum Poir.(2,64 %). Este sitio tiene la particularidad de ser el que presenta la mayor distancia entre rodales lo cual provocó la similitud con los campos naturales de Bañados de Medina y de Paso Bonilla.
En el grupo 2 se asocian a menor distancia los rodales de E. globulus y P. taeda con una orientación este-oeste (MEggE y MPtE). Esta mayor similitud se debe principalmente a los TP MCS y F, (0,15 y 0,43 %, respectivamente). El TP MCS relevado en el sitio MEggE Baccharis trimera (Less.) DC (1,06 %), mientras que en el sitio MPtE fue Erechthites hieraciifolia (L.) Raf. (1,20%). Estos sitios presentan una diferencia mayor en el TP T,(68,45 y 60,78%, MEggE y MPtE respectivamente) y donde las especies más asociadas a E. globulus (MEggE) fueron A. fissifolius (22,19 %), P. notatum (21,69 %), Dichanthelium sabulorum (Lam.) Gould & C.A. Clark (Panicum sabulorum Lam.) (11,59 %) y Piptochaetium montevidense (Spreng.) Parodi (6,58 %) y a P. taeda (MPtE) Piptochaetium bicolor (Vahl.) E. Desv.(24,59 %), P. montevidense (16,81 %) y Andropogon ternatus (Spreng.) Nees (8,51 %). Luego se agrupan los sitios CcSE y SCNN por la similitud en los TP F y D principalmente. La única especie de TP F para el sitio CcSE fue P. dilatatum (1,89 %) y para el sitio SCNN P. dilatatum (1,21 %) y Digitaria aequiglumis (Hack. & Arechav.) Parodi (0,2 0%). Las diferencias encontradas para los restantes TP no superan el 5,55 %. A este subgrupo se asocia el sitio MEggN, debido a la similitud en los TP T y D (74,97 y 0,40 %). Las especies más importantes de TP T fueron A. fissifolius (23,05 %), P. notatum (22,62 %), P. montevidense (8,68 %), S. setosa (5,13 %), S. hians (3,80 %) y Paspalum pumilum Nees (3,70 %) y para el TP D Sporobolus indicus (L.) R. Br.(0,30 %). La mayor diferencia en promedio se dio en el TP O (10,04% a favor del los sitios CcSE y SCNN). Por último a una distancia mayor se asocia el sitio SCNE, por la cobertura del TP F constituido por P. dilatatum (6,27 %) y D. aequiglumis (1,77 %), con el máximo valor relevado en este estudio y por la ausencia de MCS.
En el grupo 3 se asocian en primera instancia los callejones cortafuego con un ancho 4,66 m de los rodales de P. taeda (CcPN y CcPE), debido a la similitud en cobertura de TP T (diferencia de 0,07 %), siendo las principales especies P. montevidense (13,85 y 9,54 %), P. bicolor (8,87 y 8,68 %), P. notatum (3,81 y 5,74 %), A. fissifolius (3,46 y 5,63 %) y S. hians (3,21 y 5,88 %). Es de destacar la baja presencia de A. fissifolius y P. notatum en este ambiente menos iluminado lo cual es coincidente con lo reportado para P. notatum por Zarza24 y Berreta42. Asimismo se aprecia un cambio en el tipo funcional (metabolismo C3 vs C4), con un aumento de las especies T C3 P. montevidense y P. bicolor), lo cual es debido al menor requerimiento de ATP en el ciclo C34)(43. Para los restantes TP (MCS, ME/MM, D y O) las diferencias no superaron el 4,42 %. A una mayor distancia se agrupan los sitios MBEgN y CcBE, por la menor diferencia en el TP MCS (0 y 2,27 %) y por la ausencia de TP F. Las especies MCS solamente se relevaron en el sitio CcBE y fueron Acanthostyles buniifolium (Hook. ex Arn.) R.M. King & H. Rob.(Eupatorium buniifolium (Hook.) ex Arn.) (1,86 %), Senecio selloi (Spreng.) DC (0,26 %) y B. trimera (0,15 %). La diferencia más importante fue en el TP T, con una cobertura de 17,08 % en el ambiente menos iluminado y de 33,25 % en el callejón cortafuego. Este comportamiento podría ser explicado por los niveles de iluminación contrastantes, que provoca una mayor riqueza especifica y un mayor porcentaje de cobertura de especies tiernas que duplicó al del ambiente más sombreado (MBEgN 5 especies con una cobertura de 16,18 % y CcBE 15 especies con una cobertura de 33,25 %). Por último se asocia el sitio MPtN, por menor diferencia en la cobertura de TP D (0,73 %) y por la ausencia de TP F.
En última instancia se conforma el grupo 4 formado por un callejón cortafuego (CcBN), cuya particularidad es el valor máximo en cobertura del TP O (63,99 %). En esta se destacan las especies Carex sororia Kunth (54,62 %), Cyperus meridionalis Barros (3,91%) y otras Ciperáceas (3,15 %).
En la Figura 5 se presenta el diagrama Biplot correspondiente al análisis de componentes principales. Los CP1 y CP2 resumen el 60,8 % de la variabilidad ocurrida al clasificar por tipos productivos (TP) la vegetación desarrollada bajo rodales, callejones cortafuego y campo natural.
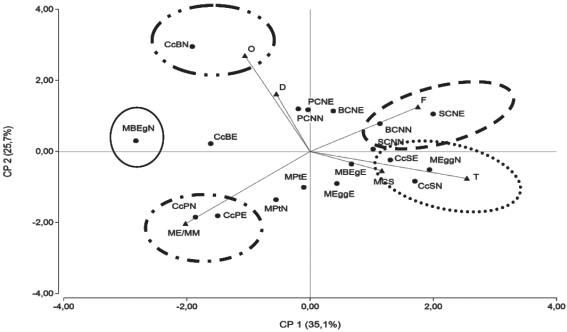
Figura 5: Biplot correspondiente al análisis de componentes principales basado en la cobertura de los tipos productivos (▲) según sitio (•).
Como se puede apreciar en la Figura 5 se generan algunos grupos definidos, mientras que otros sitios se disponen en posiciones intermedias. Sobre el CP1 a valores positivos las transectas CcSE, CcSN y MEggN presentan una mayor tendencia hacia TP T, esta variable se asoció positivamente con el CP1 y negativamente con el CP2 (0,63; -0,19). Sobre el CP1 y hacia valores positivos los sitios BCNN y SCNE presentan mayor asociación con los TP F, asociándose positivamente con el CP1 y el CP2 (0,43; 0,31). Se ubican en situaciones intermedias los sitios BCNE con una mayor proporción de especies de tipo productivo D y O; y SCNN con una proporción similar de especies F y T, pero con una mayor proporción de ME/MM y MCS que los sitios antes mencionados.
Por otra parte sobre el CP2 y hacia valores negativos las transectas CcPN y CcPE presentan una clara asociación con ME/MM, lo cual puede estar relacionado a los anchos de los callejones cortafuego (4,66 m), generándose un ambiente con mayores restricciones lumínicas y donde exhiben una adaptación Hypoxis decumbens L., D. microcalyx, Kyllinga odorata Vahl. (Cyperus sesquiflorus (Torr.) Mattf. & Kük.) y J. capillaceus.
En última instancia sobre el CP2 y hacia valores positivos de los componentes TP la transecta CcBN presentó una mayor correspondencia con tipos productivos O, lo cual es explicado por el alto porcentaje de especies de este TP relevadas (63,99 %).
Valor Pastoral Corregido (VPc)
La caracterización de los sitios de muestreo por el VPc permitió diferenciar dos grupos nítidos de sitios (Figura 6). Los agrupamientos fueron testados no rechazándose el número de grupos previsto siendo el p-valor: 0,285.
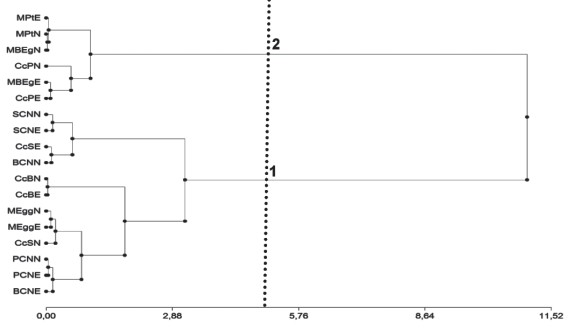
Figura 6: Dendrograma de sitios caracterizadas por el Valor Pastoral Corregido. Se diferencian dos grupos nítidos con un P-valor = 0,285.
El grupo 1 presentó un VPc promedio de 4,22 (DE ± 1,06) y el grupo 2 correspondientes a plantaciones forestales de E. grandis, las plantaciones forestales y callejones cortafuego de P. taeda, con un VPc promedio de 0,69 (DE = 0,59).
En el grupo 1 se agruparon doce sitios, donde se relevaron un total de 150 especies, lo cual representa un 87,2 % del total de especies. Dentro de este grupo podemos identificar dos sub grupos, el primero comprende a ocho sitios en los cuales se relevaron 141 especies (82 %del total de especies relevadas), de las cuales las más importantes son Andropogon lateralis Nees, A. argentinus, A. fissifolius, C. meridionalis, C. sororia, M. selloana, C. dactylon, K. odorata, Cyperus sp., Fimbristylis spadicea (L.) Vahl, S. hians, P. sabulorum, P. dilatatum, P. notatum, P. pumilum, P. montevidense, P. stipoides, S. setosa, N. charruana, Jarava plumosa (Spreng.) S.L.W. Jacobs & J. Everett (Stipa papposa Nees) y N. neesiana.
El segundo subgrupo está integrado por seis sitios relevándose 81 especies (47,1 % del total de especies relevadas) siendo las principales especies que afectan al VPc A. fissifolius, C. sororia, M. selloana, C. dactylon, C. meridionalis, K. odorata, Cyperus sp., D. aequiglumis, D. sabulorum, Eragrostis bahiensis Schrad. ex Schult., F. spadicea, S. hians, P. dilatatum, P. notatum, Paspalum plicatulum Michx., P. pumilum, P. montevidense, P. stipoides, S. setosa, N. charruana, J. plumosa y N. neesiana. Este grupo (doce sitios) presentó valores de cubierta vegetal que oscilaron entre 60,47 % (rodal de E. globulus) y 98,30 %(campo natural de Paso Bonilla), debido a que incluye desde rodales de E. globulus, callejones cortafuego (Bañados de Medina y Sarandí del Quebracho) y campos naturales de las localidades consideradas en este estudio.
El grupo 1 se caracteriza por compartir dos especies entre los 12 sitios, S. hians y J. capillaceus, lo cual corresponde al 1,33 % de las especies relevadas, pero si se excluye los sitios de campo natural de Paso Bonilla el número de especies en común asciende a 5, esto puede estar explicado por la mayor diferencia ecológica y ambiental que opera en estos sitios (PCNN y PCNE). El VPc de 4,22 obtenido para este agrupamiento está indicando que la cría de vacunos sería el destino más adecuado para estas comunidades.
El grupo 2 lo integraron seis sitios, donde se relevaron 103 especies, lo cual representa un 59,9 % del total de especies y una cubierta vegetal muy variable que osciló entre 6,1 % y 53,2 %, correspondiendo a un rodal y un callejón cortafuego de P. taeda con orientación N-S respectivamente, se observan cambios en la composición botánica que no modifican el VPc.
Solo H. decumbens L. y D. microcalix son comunes a los seis sitios (1,94 % del total), por otro lado debemos enfatizar que el principal aspecto que determina el bajo Valor Pastoral corregido es el área de cobertura vegetal, lo cual en estos sitios está afectada principalmente por la superficie no recubierta por vegetación y donde los RF presentan una mayor cobertura, coincidiendo esto con Mead44 el cual afirma que los árboles afectan a las pasturas entre otros factores a través de los restos forestales (acículas) depositados sobre el suelo generando una interferencia física.
La baja cobertura vegetal, sumado a bajas frecuencias de especies T y F (inferiores al 20 %) provoca marcadas diferencias en las vegetaciones resultando en una base forrajera pobre y degradada. Dado el escaso valor para la producción ganadera las plantaciones forestales de E. grandis con poblaciones de 1000 árboles ha-1, P. taeda con poblaciones de 1333 árboles ha-1 y callejones cortafuegos con anchos menores a 5 m, no deberían considerarse como área de pastoreo efectiva y no deben considerarse al ajustar la carga del sistema pastoril.
Los VP calculados para los grupos oscilan entre 5,04 (Grupo 1) y 3,92 (Grupo 2), mostrando relativa similitud, pero al corregir por la cobertura vegetal se generan las diferencias observadas entre los agrupamientos; reflejando el peso de la cobertura vegetal sobre la aptitud pastoril de las comunidades.
Conclusiones
Existen diferencias en la cobertura del suelo entre los sitios relevados, las comunidades vegetales desarrolladas bajo plantaciones forestales de E. grandis, y P. taeda presentaron la menor cobertura vegetal. En los callejones cortafuego la cobertura vegetal es dependiente de las distancias entre rodales, dónde, las menores distancias provocan una reducción del área ocupada por la vegetación herbácea. Todas las comunidades presentan valores similares en el valor pastoral; cuando introducimos el área de cobertura vegetal para la estimación del VPc, se generan diferencias, indicando la importancia de la cobertura vegetal en la determinación de la aptitud pastoril de una comunidad herbácea, por lo que, comunidades desarrolladas bajo callejones cortafuego y rodales de P. taeda y de E. grandis presentaron una inferior aptitud ganadera al resto de las situaciones.

