











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo

 Permalink
PermalinkIntroducción
Phytophthora sojae Kaufmann & Gerdemann es un Oomicete del reino Stramenopila que causa la podredumbre de raíz y de base de tallo en soja. Este patógeno puede infectar a la soja en cualquier estadio fenológico, provocando damping-off o muerte de plántulas en pre y pos emergencia, y la infección de la raíz y del tallo de plantas adultas. En 1998, se estima que se perdieron 1,27 millones de toneladas a causa de esta enfermedad en los cinco países de mayor producción de soja a nivel mundial (Dorrance y Grünwald, 2009). En EEUU, dependiendo del año, es clasificada como la segunda o tercera enfermedad más importante de la soja (Wrather y Koenning, 2009). Wilcox y St. Martin (1998) registraron mermas en la producción de grano de entre 65 % y 93 %, comparando el rendimiento de isolíneas de sojas susceptibles y resistentes a esta enfermedad. En Argentina se han reportado incidencias del 60 y 70 % en Pergamino y Marcos Juárez, respectivamente (Vallone et al., 1999). En Brasil se reportó por primera vez en Río Grande del Sur en la zafra 1994-1995 (Costamilan et al., 1996). No fue importante hasta la zafra 2005-2006, donde se produjeron importantes fallas en la implantación en suelos compactados, lo cual conllevó a la resiembra de grandes extensiones. En Uruguay, hasta el momento, no había sido reportada en el cultivo de soja.
En los últimos 50 años, a nivel mundial, esta enfermedad se ha manejado mediante variedades de soja con genes de resistencia mayores o Rps (Grau et al., 2004; Schmitthenner, 1985). Se conocen actualmente 17 genes (11 loci y una serie alélica en dos de ellos) en soja que confieren resistencia a P. sojae (Grau et al., 2004; Lin et al., 2013; Sun et al., 2010; Zhang et al., 2013). Estos genes funcionan de acuerdo al postulado de gen-a-gen propuesto por Flor (1955). Este postulado propone que para cada gen de resistencia en la planta existe un correspondiente gen de avirulencia en el patógeno. Este último codifica un elicitor que directa o indirectamente interactúa con el producto del correspondiente gen de resistencia en la planta, causando la reacción de resistencia o hipersensibilidad (Parker, 2009).
Más de 200 patotipos del patógeno han sido reportados mundialmente en soja (Dorrance, McClure y deSilva, 2003), lo que sugiere una alta variación genética con respecto a la virulencia en la población natural. Los patotipos se determinan utilizando un set de 8 a 14 diferenciales de soja, donde cada línea de soja posee un gen de resistencia o Rps. Hasta el año 2000 se habían descrito 55 razas, pero debido al surgimiento continuo de nuevas combinaciones de genes de virulencia en el patógeno, la asignación de un número a la raza se tornó indescifrable (Grau et al., 2004). Por tal motivo, actualmente se describe el patotipo utilizando directamente la fórmula de virulencia, o sea indicando el gen Rps de la soja a la cual el aislado es capaz de infectar.
Las variedades de soja sembradas en el país generalmente se comercializan con la información del criadero indicando la resistencia a distintas razas de P. sojae. Es fundamental conocer qué razas o patotipos predominan en las condiciones de Uruguay para determinar su compatibilidad con las variedades sembradas y, a su vez, para seleccionar por resistencia dentro de un programa de mejoramiento de soja. El objetivo de este trabajo fue reportar por primera vez a Phytophthora sojae afectando soja en Uruguay y determinar los patotipos de un número limitado de aislados.
Materiales y métodos
Aislamiento y purificación del patógeno
Plantas con síntomas de podredumbre de raíz y tallo, en estadios entre dos nudos y plena floración, de las variedades A 5009 RG, A 5509 RG, DM 6.2i, DM 7.0i y Cardinal GE 590 ci de las localidades de San Pedro, Semillero y Manantiales del departamento de Colonia y Trinidad en el departamento de Flores, de las zafras 2013-2014 y 2014-2015, fueron utilizadas para realizar los aislamientos. Las mismas se lavaron y se dejaron debajo de agua corriendo por 12-16 horas, luego se cortó y esterilizó la lesión con hipoclorito al 1 %, luego con alcohol al 70 % y se enjuagó con H2O estéril. Se cortó una pequeña porción entre la zona afectada y parte sana y se colocó debajo del medio de cultivo V8 diluido (40 ml de jugo de vegetales V8, 0,6 g CaCO3, 0,2 g extracto de levadura, 1 g sucrosa, 20 g agar, 1000 ml de dH2O). Los aislados logrados fueron purificados utilizando la técnica de punta de hifa (French y Herbert, 1980) y guardados en trozos de medio de cultivo V8 diluido en agua estéril a temperatura ambiente (Schmitthenner y Bhat, 1994).
Identificación del patógeno
La identificación fue realizada observando las características de crecimiento y la morfología de las estructuras en medio de cultivo V8 diluido y luego de sucesivos lavados en dH2O estéril a 10 °C. Para la identificación molecular, el ADN genómico fue extraído del micelio de tres aislados puros molidos con nitrógeno liquido por método CTAB (Innis et al., 1990). Los tres aislados seleccionados para la identificación molecular (Ps04, Ps06 y Ps11) fueron trabajados por duplicado y seleccionados en base a cantidad/ calidad del ADN extraído. La calidad y la concentración del ADN fueron determinadas por electroforesis en agarosa al 0,8 % y utilizando un espectrofotómetro NanoDrop 2000 (Thermo Fisher Scientic, USA) a 260 nm. El ADN fue amplificado por PCR convencional utilizando los cebadores ITS 5 (52 -GGAAGTAAAAGTCGTAACAAGG) e ITS 4 (52 -TCCTCCGCTTATTGATATGC) (White et al., 1990). Las condiciones de ciclado se realizaron siguiendo el protocolo descrito por Ristaino et al. (1998). El largo del amplicon fue testeado en 1,5 % gel de agarosa. Los productos del PCR fueron secuenciados en ambos sentidos (forward y reverse) utilizando el servicio estándar de Macrogen Inc, Korea del Sur (http://www.macrogen.com). Las secuencias forward y reverse, así como la secuencia consenso, fueron editadas utilizando el software BioEdit v7.2.5 (http://www.mbio.ncsu.edu/bioedit/bioedit.html). Ambas secuencias fueron editadas eliminando los extremos con mala calidad de secuencia. Posteriormente ambas secuencias fueron alineadas, y la secuencia consenso fue sometida a búsqueda BLAST disponible en la base de datos de NCBI (http://blast.ncbi.nlm.nih.gov) y depositadas en GenBank.
Determinación de patotipos
Los patotipos fueron determinados utilizando la técnica de inoculación en hipocótilo en 14 líneas diferenciales de soja (Dorrance et al., 2008; Schmitthenner y Bhat, 1994). Los diferenciales utilizados fueron: Harlon (Rps 1a, Blackhawk source), L77-1863 (Rps 1b), Williams 79 (Rps 1c), Wu An - PI 103091 (Rps 1d), Williams 82 (Rps 1k), L82-1449 (Rps 2), L83-570 (Rps 3a), L91- 8347 (Rps 3b), L92-7857 (Rps 3c), L85-2352 (Rps 4), L85-3059 (Rps 5), L89-1581 (Rps 6), L93-3258 (Rps 7), y Williams (susceptible universal). La soja se sembró en un sustrato mezcla (5:1:1 tierra:turba:vermiculita) en bandejas en invernáculo. A los 7-10 días de la siembra, 10 plántulas de cada diferencial fueron inoculadas efectuando una incisión, tipo ojal, en el hipocótilo con la aguja de una jeringa hipodérmica. Dentro de la lesión, utilizando la jeringa, se virtieron 0,2 a 0,4 ml de macerado del patógeno con 7-10 días de crecimiento en medio de cultivo V8 diluido (Dorrance et al., 2008). Las bandejas se cubrieron con nylon por 16-24 h para prevenir secado. Siete a diez días pos inoculación se registró la incidencia de plántulas muertas o que mostraron lesiones necróticas expandidas. Un diferencial es considerado susceptible si al menos el 70 % de las plántulas muestran esta sintomatología. El test se realizó dos veces para cada aislado.
Resultados
Se obtuvieron 14 aislados purificados de Phytophthora sojae de las plantas sintomáticas (Cuadro 1). Las colonias fueron identificadas como tales por presentar márgenes algodonosos densos, con oosporas de paredes gruesas de 20-42 μm de diámetro sumergidas en el medio, con anteridios de tipo parágino y anfígino. En el agua estéril se desarrollaron esporangios no papilados, no caducos, ovoides u obpiriformes de 24-68 x 15-28 μm, que liberan zoosporas biflageladas arriñonadas. Los esporangióforos presentaron proliferación interna y encadenada o terminal. Las características evaluadas coinciden con el grupo V de Waterhouse, con el clado 7 de Kroon y con la clasificación taxonomía general del patógeno (Dorrance et al., 2007; Erwin, Bartnicki-Garcia y Tsao, 1983; Erwin y Ribeiro, 1996; Hartman et al., 2015; Kroon et al., 2012; Waterhouse, 1963). Las secuencias obtenidas de la región ITS de los tres aislados analizados presentaron 99 % o 100 % de similitud con la especie P. sojae en la base de datos consultada (NCBI) (Cuadro 2).
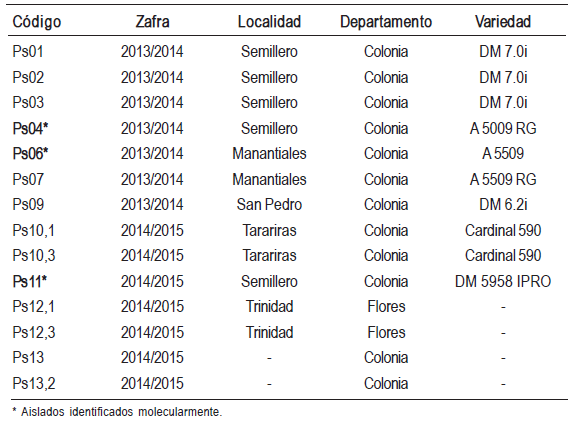
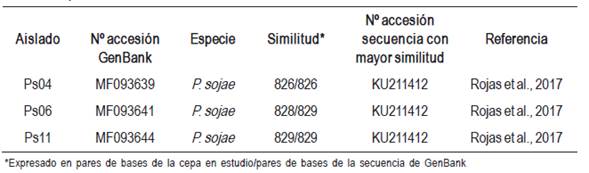
Cuadro 2: Análisis de similitud de las secuencias ITS de los aislados en estudio, con secuencias de referencias publicadas en GenBank.
Se determinaron siete patotipos diferentes de los 14 aislados de P. sojae estudiados. El 57 % de los aislados pertenecieron a un único patotipo; vir 1b, 3a, 3b, 5 (sin nombre de raza conocido). Los seis aislados restantes pertenecieron cada uno a un patotipo diferente, tres de ellos con nombres de razas conocidas: raza 4, 14 y 25 (Cuadro 3).
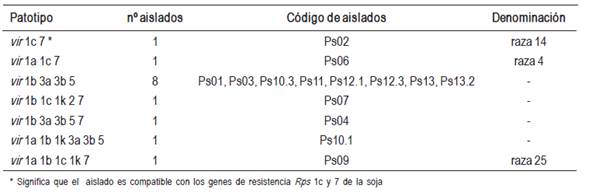
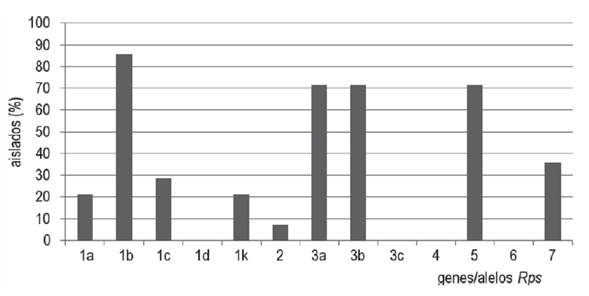
Nueve de los 13 genes de resistencia de la soja estudiados fueron compatibles con al menos un aislado de P. sojae (Rps 1a, 1b, 1c, 1k, 2, 3a, 3b, 5 y 7) (Figura 1). Más del 70 % de los aislados fueron compatibles con los genes Rps 1b, 3a, 3b y 5 (Figura 1). Contrariamente, los diferenciales de soja que poseen los genes Rps 1d, 3c, 4 y 6 mostraron reacciones de resistencia al 100 % de los aislados estudiados.
Discusión
Los caracteres morfológicos y los análisis moleculares realizados permiten confirmar la identidad del patógeno como Phytophthora sojae, siendo este el primer reporte de P. sojae en Uruguay. Desde que se encontró la podredumbre de raíz y base de tallo de la soja, sólo se han visto afectadas plantas aisladas en el cultivo (Stewart, com pers). Contrariamente, la fase de damping-off o muerte de plántulas ha producido importantes fallas de implantación (Stewart, com pers.), que al igual que en Brasil han determinado la resiembra de grandes extensiones (Costamilan et al., 1996).
A pesar del número limitado de aislados del trabajo, la variabilidad de patotipos encontrada fue mayor a la esperada para este nuevo patógeno. Generalmente cuando la enfermedad es recientemente introducida en una región o país, se encuentra una raza predominante. En EEUU y en Canadá prevaleció la raza 1 (vir 7) hasta la década del 90 (Anderson y Buzzell, 1992; Laviolette y Athow, 1981; Schmitthenner, Hobe y Bhat, 1994). Tanto en Argentina como en Brasil, los primeros aislados de P. sojae encontrados en la década del 90 pertenecieron exclusivamente a la raza 1 (Barreto, Stegman de Gurfinkel y Fortugno, 1995; Costamilan y Bonato, 1996). Aparecieron nuevos patotipos a medida que la enfermedad adquirió importancia en el cultivo y se fueron liberando variedades con genes Rps. Es así que en la zafra 2013-2014 en el sudeste bonaerense se determinaron 12 patotipos distintos en 18 aislados, ninguno de los cuales correspondió a la raza 1 (Grijalba et al., 2015b). En nuestro trabajo, si bien se encontró un patotipo predominante, este no fue la raza 1, y al igual que en el sudeste bonaerense ninguno de los aislados correspondió a esta raza.
Los genes Rps comúnmente incorporados a las variedades comerciales en la región y en el mundo son Rps1a, Rps 1c y Rps1k (Grijalba et al., 2015a; Schmitthenner, Hobe y Bhat, 1994), los cuales serían efectivos para más del 70 % de los patotipos encontrados en este estudio. Estos resultados sugieren que la resistencia a la enfermedad utilizando estos genes mayores todavía es efectiva para nuestras condiciones, si bien otros genes como Rps1d, Rps 3c, Rps 4 y Rps 6 podrían ser hasta un 100 % efectivos.
El patotipo predominante vir 1b 3a 3b 5 debería ser utilizado para determinar su compatibilidad con las variedades Figura 1. Porcentaje de aislados compatibles con los genes de resistencia de la soja (Rps). de soja sembradas localmente y con el material avanzado del programa de mejoramiento de soja local, ya que este patotipo no se utiliza para seleccionar soja en los programas de mejoramiento de la región.
A nivel mundial la tendencia es al incremento de la complejidad de la virulencia de los aislados (Dorrance, McClure y deSilva, 2003; Grijalba et al., 2015b; Schmitthenner, Hobe y Bhat, 1994), que conjuntamente con la variabilidad encontrada en este estudio realza la importancia de rutinariamente hacer un seguimiento de los patotipos y de la efectividad de los genes de resistencia.

