










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Links relacionados
Compartir

 Permalink
PermalinkAgrociencia (Uruguay)
versión impresa ISSN 1510-0839versión On-line ISSN 2301-1548
Agrociencia Uruguay vol.20 no.1 Montevideo jun. 2016
Estudio preliminar de la prolificidad y habilidad materna de seis biotipos ovinos en Uruguay
Banchero Georgget1, Vázquez Andrés2, Irarí Natalia1, Ciappesoni Gabriel2, Quintans Graciela3
1Instituto Nacional de Investigación Agropecuaria La Estanzuela. Ruta 50 km 11, Colonia, Uruguay. Correo electrónico: gbanchero@inia.org.uy
2Instituto Nacional de Investigación Agropecuaria Las Brujas. Ruta 48 km 10, Canelones, Uruguay.
3Instituto Nacional de Investigación Agropecuaria Treinta y Tres. Ruta 8 km 282, Treinta y Tres, Uruguay.
Recibido: 3/10/14 Aceptado: 16/12/15
Resumen
Los resultados de las primeras evaluaciones del Proyecto Ovejas Prolíficas de INIA (2008-2011) muestran que la cruza Frisona Milchschaf x Corriedale o Finnish Landrace x Corriedale desteta 60 % más corderos que la Corriedale pura y, por otro lado, la cruza Finnish Landrace x Frisona Milchschaf desteta 85 % más que la Corriedale pura. Sin embargo y bajo las mismas condiciones productivas, no se conoce el comportamiento al parto y la producción de calostro de estos nuevos biotipos. Para recabar información, se realizó un experimento en INIA La Estanzuela, Colonia, Uruguay (34°S; 57°O) donde se utilizaron 61 borregas de 4-6 dientes nacidas en la primavera del año 2010. Se utilizaron dos razas puras: Corriedale (C.C) y Frisona Milchschaf (M.M) y las cruzas Finnish Landrace x Corriedale (F.C), Frisona Milchschaf x Corriedale (M.C), 3/4 Frisona Milchschaf - 1/4 Finnish Landrace (M.(F.M)) y 7/8 Finnish Landrace - 1/8 Corriedale (F.(FxF.C)), apareadas con carneros Texel. El incremento de sangre Finnish en la cruza redundó en un aumento en la prolificidad (2,2; 2,3 y 2,4 para 1/4; 1/2 y 7/8 Finnish) con corderos más livianos y menor asistencia al parto. Tanto la media sangre Milchschaf como la pura tuvieron una prolificidad similar e intermedia (1,8 M.C y 1,7 M.M) mientras que la asistencia al parto fue alta. La producción y composición de calostro fue similar entre biotipos a excepción de M.M que produjo más. No existió diferencia en eficiencia placentaria entre biotipos. El peso vivo al nacimiento de los corderos influyó sobre el comportamiento de los mismos en su primera hora de vida, precisamente en el tiempo desde el nacimiento hasta que lograron pararse y mamar.
Palabras clave: CARGA FETAL, COMPORTAMIENTO MATERNAL, LARGO DE PARTO, PRODUCCIÓN DE CALOSTRO, PLACENTA
Summary
Preliminary Study of Prolificacy and Maternal Ability on Six Sheep Biotypes in Uruguay
The results of the first evaluation of the Prolific Sheep Project of INIA (2008-2011) demonstrated that the crossbred biotypes Corriedale by East Friesian or Corriedale by Finnish Landrace weaned 60 % more lambs than pure Corriedale and, on the other hand, the crossbred East Friesian by Finnish Landrace weaned 85 % more than pure Corriedale. However, and under the same productive conditions, the behavior at lambing and colostrum production of these new biotypes are unknown. In order to find out, an experiment was conducted at INIA La Estanzuela, Colonia, Uruguay (34°S; 57°O) where 61 female lambs 4-6 teeth born in the spring of 2010 were evaluated. Two pure breeds were used: Corriedale (C.C) and East Friesian (M.M), and the crosses Finnish Landrace x Corriedale (F.C), East Friesian x Corriedale (M.C), 3/4 East Friesian - 1/4 Finnish Landrace (M.(F.M)), 7/8 Finnish Landrace - 1/8 Corriedale (F.(FxF.C)), mated with Texel rams. The increase in Finnish blood resulted in more prolificacy (2.2; 2.3 and 2.4 for 1/4; 1/2 and 7/8 Finnish with lighter lambs and less lambing assistance. Both half and pure East Friesian had similar and moderate prolificacy (1.8 M.C and 1.7 M.M) whereas lambing assistance was high. Colostrum production and composition was similar between biotypes, with the exception of M.M, that produced more. There were no differences in placental efficiency between biotypes. Birth weight of lambs affected their behavior in the first hour of life, specifically the time from birth to first stand up to suckle.
Keywords: FETAL LOAD, MATERNAL BEHAVIOR DURATION OF DELIVERY, COLOSTRUM PRODUCTION, PLACENTA
Introducción
Los parámetros productivos de mayor incidencia sobre el resultado económico de los sistemas ovinos semi-intensivos e intensivos son el comportamiento reproductivo y la habilidad materna de las ovejas (Ganzábal, 2013). Las evaluaciones realizadas dentro del proyecto de transferencia integral del Secretariado Uruguayo de la Lana (SUL) demostraron que con un destete del 90 % en razas laneras se puede alcanzar una producción de carne de 190 kg por hectárea (SUL, 2006). Si realizamos cruzamientos terminales con carneros de razas carniceras sobre las mismas ovejas, la producción de carne se puede incrementar un 26 % (Montossi et al., 2013). Asimismo, si cambiamos la madre por un biotipo prolífico, la producción de carne puede duplicarse con la misma producción de lana (Montossi et al., 2013). De este modo, utilizando biotipos prolíficos como madres y razas carniceras como padres, se pueden obtener destetes de 150 % y corderos pesados de 35-38 kg que son faenados con seis meses de edad, lo que implica una producción de carne anual de 236 a 370 kg/ha y 50 kg/ha de lana (Montossi et al., 2013).
En efecto, las evaluaciones realizadas dentro del proyecto Prolíficas de INIA (2008-2012) muestran que las madres puras Corriedale (C.C), Frisona Milchschaf (M.M) y Finnish Landrace (F.F) tienen el potencial de destetar 125, 159 y 206 %, mientras que las madres cruza Frisona Milchschaf x Corriedale (M.C), Finnish Landrace x Corriedale (F.C) y Finnish Landrace x Frisona Milchschaf (F.M) tienen el potencial de destetar 148, 178 y 205 % respectivamente (Ciappesoni et al., 2014). En dicho proyecto, las medidas al parto fueron predefinidas e incluyeron solamente dos recorridas diarias para identificar los corderos nacidos/muertos. Por lo tanto, no fue posible evaluar el comportamiento al parto, tanto en producción de calostro como en comportamiento maternal y neonatal de los corderos, características que escapaban al objeto de estudio.
Para este trabajo se utilizaron los biotipos del estudio antes mencionado, sustituyendo la raza F.F debido a su escasa presencia en el país y su falta de adaptación por animales que se fueron absorbiendo sobre base Corriedale (7/8Finnish Landrace y 1/8 Corriedale, F.(FxF.C)), y la F.M por uno de los biotipos que sus predicciones sugerían como promisorios (3/4 Frisona Milchschaf y 1/4 Finnish Landrace, Mx(F.M)).
El objetivo establecido fue evaluar algunas características relacionadas al desempeño reproductivo en seis biotipos ovinos, de modo de mejorar nuestra comprensión acerca de los factores que pueden estar impactando en los resultados globales del proyecto.
Materiales y métodos
Los materiales y métodos generales utilizados en este trabajo son presentados en el artículo de Banchero et al. (2014), «Estudio preliminar del crecimiento, desarrollo e indicadores reproductivos de hembras de seis biotipos ovinos en Uruguay». Los animales utilizados y los materiales y métodos particulares son presentados a continuación. En el presente artículo solo se tomaron en cuenta las ovejas gestantes al momento del diagnóstico de preñez y cuya gestación llegó a término. Se evaluaron dos razas puras: Corriedale (C.C; n = 10) y Frisona Milchschaf (M.M; n = 10) y las siguientes cruzas: Finnish Landrace x Corriedale (F.C; n = 10), Frisona Milchschaf x Corriedale (M.C; n=14), 3/4 Frisona Milchschaf - 1/4 Finnish Landrace (M.(F.M); n = 9), 7/8 Finnish Landrace - 1/8 Corriedale (F.(FxF.C); n = 8). Las características reproductivas evaluadas incluyeron: prolificidad, asistencia al parto, lactogénesis (producción y calidad de calostro), duración del parto (DP), peso vivo de los corderos al nacimiento (PVN), comportamiento de los corderos en su primer hora de vida y características de la placenta.
Prolificidad y asistencia al parto
En el otoño del 2013 todas las ovejas fueron apareadas por primera vez bajo monta natural. El servicio comenzó a finales de marzo y tuvo una duración de 32 días. Durante el periodo de apareamiento todas las ovejas se manejaron juntas con tres carneros de la raza Texel, con el fin de no favorecer a determinado biotipo por asignarle carneros posiblemente superiores para algunas de las características estudiadas en la progenie. Para diagnosticar la gestación y conocer el número de fetos por oveja se realizó una ecografía a los 70 días de introducidos los carneros con un ecógrafo Aloka 500 (Tokio, Japón) y un transductor de 7.5 MHz.
Todas las ovejas se manejaron en forma conjunta en condiciones pastoriles sin suplementación. Las pasturas utilizadas fueron pasturas artificiales (alfalfa y trébol rojo; 11 a 27 % de PC y 24 a 46 % de FDN) y verdeos (raigrás; 9 a 13 % PC y 25 a 47 % de FDN), con una asignación de forraje mínima del 6 % del peso vivo. Diez días previo a la primera fecha probable de parto, todas las ovejas gestantes se manejaron en un potrero de 2 ha donde se les suministró 1 kg de ración totalmente mezclada (87,7 % MS, 17,4 % PC; 2.46 MCal de EM/kg MS) por animal y por día. Además contaban con fardo de alfalfa ad libitum (87,7 % MS, 17 % PC y 2,3 MCal de EM/kg MS) para cubrir los requerimientos preparto (MAFF, 1975).
Al momento del parto se registró en las ovejas el número de corderos nacidos vivos o muertos y la asistencia al parto. En las ovejas que tuvieron gestaciones simples se registró la necesidad de asistencia al parto o no, mientras que en las ovejas con gestaciones múltiples se tomaron como asistidas aquellas ovejas que precisaron ayuda en el parto de al menos uno de sus corderos. La decisión de asistencia al parto fue tomada cuando hubo evidencia de mala presentación del cordero o cuando no había progreso del parto en un cordero normalmente presentado, transcurrida una hora luego de visualizarse cualquier miembro o cabeza en la vulva de la oveja (Dwyer y Lawrence, 1999).
Lactogénesis
Al momento del parto se registró el volumen de la ubre, para su cálculo se tomaron las medidas latero-lateral y antero-posterior de la ubre, previo a la realización del ordeñe (Bencini y Purvis, 1990). Una hora después de nacido el último cordero, las ovejas fueron inyectadas con 5 UI de oxitocina (técnica de Doney et al., 1979), se ordeñó a mano completamente un pezón, y se registró peso y viscosidad a través de un escore de Mc Cance y Alexander (1959). La producción total se estimó como la producción obtenida en dicho ordeñe x 2. Se extrajo una muestra de calostro (aproximadamente 20 ml), la cual se preservó con Lactopol® (2-Bromo-2-Nitropropano-1,3-diol) y almacenó a -15 °C hasta el análisis. La composición del calostro fue analizada en el laboratorio de calidad de leche de INIA a través de un Lactoscan, Milkanalyzer (Nova Zagora, Bulgaria), se determinaron los porcentajes de grasa, proteína, lactosa y sólidos no grasos.
Duración del parto, peso y comportamiento de los corderos en su primera hora de vida
En cada uno de los corderos se registró: i) la duración de la fase II del parto, desde la aparición de las pezuñas hasta la expulsión total del cordero (Grunert et al., 1971); ii) peso al nacer; y iii) comportamiento en la primer hora de vida registrándose los tiempos en que intenta pararse (IP), logra pararse (LP), intenta mamar (IM) y logra mamar (LM). Se considera intento cuando el animal no logra superar los seis segundos de pie, y éxito cuando el cordero se mantiene de pie por al menos seis segundos (Murphy, 1999).
Características de la placenta
Se pesaron las placentas y se registró el número y peso de los cotiledones, se estimó la eficiencia placentaria como gramos de cordero producido por cada gramo de placenta, y la eficiencia cotiledonaria como gramos de cordero producido por cada gramo de cotiledón.
Análisis estadístico
a) Prolificidad y asistencia al parto
El peso al inicio del servicio (20 de marzo del 2013) se analizó mediante un modelo lineal y el procedimiento GLM de SAS (Statistical Analysis System, Version 9.2, 2008). Se utilizó un modelo corregido por el efecto fijo del biotipo de la oveja y además se incluyó el tipo de nacimiento y la edad al servicio. El análisis estadístico de prolificidad se realizó a través del estudio del número de corderos nacidos vivos por oveja parida (TP: 1, 2 o 3). Se analizó asumiendo una distribución multinomial (ordenada) y función de vínculo Logit acumulado. Se modeló la probabilidad de tener una mayor prolificidad. Además de corregirse por biotipo como efecto fijo, también se corrigió por los días desde una fecha fija (pesada preparto) hasta el parto como covariable, con el objetivo de corregir las variaciones estacionales existentes durante el periodo de apareamiento (por ejemplo variaciones sobre la tasa ovulatoria y por consiguiente sobre la prolificidad). El análisis se realizó mediante el procedimiento GENMOD del paquete estadístico SAS (Statistical Analysis System, Version 9.2, 2008). Para evaluar la asistencia al parto por biotipo se realizaron tablas de contingencia. Para detectar diferencias significativas se realizó un test exacto de Fisher.
b) Lactogénesis
El volumen de la ubre y la producción y composición de calostro se analizó asumiendo una distribución normal mediante el procedimiento GLM del paquete estadístico SAS (Statistical Analysis System, Version 9.2, 2008). Se corrigió por los efectos fijos de biotipo y el tipo de parto (1, 2 o 3) de la oveja. También se incluyó en el modelo, como en el caso anterior, los días desde una fecha fija (pesada preparto) hasta el parto como covariable con el objetivo de corregir las variaciones estacionales existentes durante el periodo de partos. Para evaluar la viscosidad y el color del calostro se realizaron tablas de contingencia. Para detectar diferencias significativas se realizó un test exacto de Fisher.
c) Duración del parto, peso y comportamiento de los corderos en su primera hora de vida
i) El peso al nacimiento de los corderos se evaluó asumiendo una distribución normal mediante el procedimiento GLM del paquete estadístico SAS (Statistical Analysis System, Version 9.2, 2008). Se corrigió por los efectos fijos de biotipo, sexo y el tipo de nacimiento (1, 2 ó 3) del cordero
(ii, iii). La evaluación de la duración de la fase II del parto y del comportamiento de los corderos se realizó, como en el caso anterior, asumiendo una distribución normal mediante el procedimiento GLM del paquete estadístico SAS (Statistical Analysis System, Version 9.2, 2008). Se corrigió por los efectos fijos de biotipo, sexo y tipo de nacimiento (1, 2 ó 3) del cordero. Asimismo se estudió un modelo alternativo con la inclusión del peso vivo al nacimiento de los corderos como covariable.
d) Características de la placenta
Las características de la placenta fueron evaluadas asumiendo una distribución normal, con el mismo modelo que para la evaluación de lactogénesis. Se definió como no significativo los valores P > 0,05.
Resultados y discusión
Prolificidad
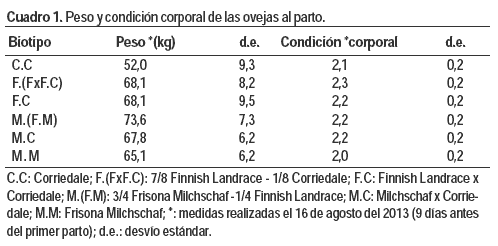
Las ovejas presentaron similar condición corporal al comienzo del servicio (2,3 ± 0,3 puntos; media ± desvío estándar). El peso vivo al inicio del servicio estuvo afectado por la edad, el tipo de nacimiento y el biotipo (P < 0,0047; P = 0,0415 y P = 0,0007 respectivamente). Las medias corregidas de los pesos al inicio del servicio y sus errores estándares fueron: C.C 51,2 ± 2,0; M.M 55,4 ± 2,0; F.C 55,6 ± 1,9; M.C 60,9 ± 1,6; M.(F.M) 61,3 ± 1,9; F.(FxF.C) 53,7 ± 2,0 kg). La prolificidad, en este caso medida como número de corderos por oveja parida, se manifestó de acuerdo a lo esperado, teniendo en cuenta las razas que componen los diferentes biotipos.
Aquellos con cierto porcentaje de Finnish tuvieron los valores más altos y duplicaron la prolificidad de la C.C (2,3 vs. 1,2 respectivamente). A su vez, a medida que el porcentaje de Finnish aumentó también lo hizo la prolificidad (2,2; 2,3 y 2,4 para 1/4; 1/2 y 7/8 proporción de sangre Finnish). Esto concuerda con lo reportado por Goot et al. (1984) quien presenta valores de 2,2; 2,5 y 3,0 corderos nacidos por oveja parida por año para 1/4; 1/2 y 3/4 proporción de sangre Finnish. Sin embargo, Lira (2007) reportó incrementos menores de prolificidad con respecto a la raza Corriedale, siendo de 1,07, 1,10 y 1,39 (fetos ecografiados por oveja preñada) para vientres C.C, 1/4 F 3/4 C y F.C. Es de destacar que estos experimentos se realizaron en condiciones con limitantes nutricionales donde seguramente no se haya podido expresar el potencial genético.
Para nuestro experimento, se encontraron diferencias significativas (P < 0,05) para los contrastes entre F.(FxF.C) - C.C; entre F.C - C.C y entre M.(F.M) - C.C. El logaritmo de la razón de oportunidades (Log RO) para la diferencia F.(FxF.C) - C.C es de 3,97, su exponencial (Exp (Log RO)) es de 53. Este valor indica que las oportunidades de que el biotipo F.(FxF.C) presente mayor prolificidad es 53 veces mayor que las oportunidades del C.C (53:1). Esto equivale a decir que la probabilidad de que la prolificidad del biotipo F.(FxF.C) sea mayor a la del C.C es de 0,982 (Probabilidad = Exp Log RO/(Exp Log RO+1)).
Analizando las diferencias en prolificidad según el tipo de gestación (Figura 1), el biotipo F.(FxF.C) tiene la mayor probabilidad de presentar gestación triple (P = 0,44) de todos los biotipos evaluados y una probabilidad de 0,53 de presentar gestación doble. Por otro lado, la probabilidad de que las ovejas C.C presenten gestación triple es muy baja (la menor de todos los biotipos evaluados; P = 0,0058), mientras que la probabilidad de presentar gestación única es la mayor de los biotipos evaluados (P = 0,83). En el contraste F.C - C.C, la probabilidad de que la prolificidad del biotipo F.C sea mayor que la del C.C es de 0,974.
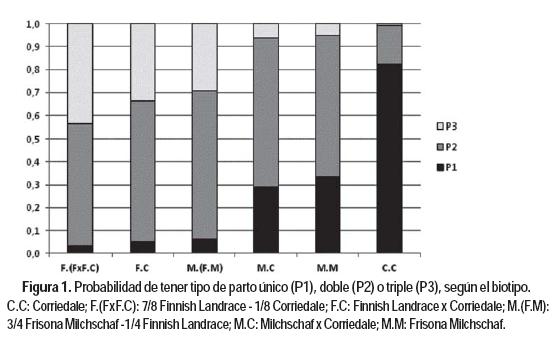
Thomson et al. (2004) en ovejas base Romney (R.R) reportan diferencias en prolificidad de 33 puntos porcentuales entre F.R y R.R a favor de las cruzas. En cuanto al contraste M.(F.M) - C.C, la probabilidad de que la prolificidad del biotipo M.(F.M) sea mayor que la del C.C es 0,963. Los biotipos M.C y M.M presentaron valores intermedios de prolificidad (1,8 y 1,7 respectivamente) no encontrándose diferencias significativas entre ellos. A pesar que las tendencias reportadas en la bibliografía son las mismas, la diferencia de prolificidad entre M.C y C.C presentada por Bianchi et al. (2000), Barbato et al. (2011) y Kremer et al. (2010) fue de sólo 26, 29 y 12 puntos porcentuales respectivamente; mientras que Thomson et al. (2004) encontraron diferencias de 18 puntos porcentuales a favor de las cruzas M.R en relación a las ovejas R.R.
El peso corporal al inicio del servicio es otro factor de gran incidencia en la determinación de la TO y por lo tanto de la prolificidad. Por ejemplo, Morley et al. (1978) reportaron un incremento de 2 % de corderos nacidos por cada kg extra a la encarnerada para Merino Australiano en Western-Australia, mientras que Ganzábal (2005) reportó un incremento de 1,7 % por cada kg vivo extra al apareamiento en Corriedale en Uruguay. Thomson et al. (2004) obtuvieron efectos significativos (P < 0,05) del peso vivo de la oveja al inicio del servicio sobre la prolificidad, reportando incrementos de 1,3, 1,1, 0,7 y 1,3 % por cada kg extra para F.R, M.R, Poll Dorset (PD).R y R.R respectivamente. En el presente experimento, el PV fue significativo sobre la prolificidad (P = 0,0114), pero aun corrigiendo por este factor, el biotipo continuó siendo significativo (P = 0,0068).
Asistencia al parto
La asistencia de las ovejas al parto varió entre los diferentes biotipos (P = 0,0001). La proporción fue mayor en las M.M y sus cruzas (M.M 50 %; M.(F.M) 44,4 %; M.C 28,6 %). En el otro extremo estuvieron las F.(FxF.C) donde ninguna oveja necesitó asistencia. Estas diferencias estarían explicadas principalmente por los pesos al nacimiento de sus corderos, ya que todas las ovejas presentaron una condición corporal al parto de 2 puntos (Cuadro 1). Los pesos al nacimiento de los corderos difirieron según el biotipo de su madre (P < 0,0001) y el tipo de nacimiento (P = 0,0007). Efectivamente, Scales et al. (1986) encontraron que la proporción de ovejas con asistencia al parto aumenta con pesos al nacer por encima de los 4,5 kg. En nuestro trabajo, la mayor diferencia de peso se presentó entre los corderos hijos de ovejas M.M con los corderos hijos de ovejas F.(FxF.C) siendo esta de 1,7 kg a favor de los primeros. Parte de esta diferencia está explicada por el tipo de nacimiento de los corderos, donde las ovejas F.(FxF.C) tuvieron mayor prolificidad que las M.M (contraste F.(FxF.C) - M.M, P = 0,0232) y los corderos de partos múltiples fueron 1,0 kg más livianos que los de parto único.
Desarrollo de la ubre, producción y calidad del calostro
El volumen de la ubre fue diferente de acuerdo al biotipo (Cuadro 2; P = 0,0042). Las ovejas M.M fueron las que presentaron mayor volumen, posiblemente por la especialización de esta raza en la producción lechera. Si comparamos el volumen de la ubre tanto lleno como vacío (volumen de la ubre - peso de calostro) entre M.M y los biotipos que no tienen sangre Milchschaf en su composición genética, vemos que las primeras presentan un volumen lleno o vacío de al menos dos veces superior a estas últimas. Sin embargo, el volumen de la ubre no fue afectado por el tipo de gestación, a diferencia de lo encontrado por Gigena y Vázquez (2005) en ovejas Corriedale, donde ovejas gestando mellizos tuvieron ubres más grandes (casi 30 % superior) al momento del parto, en relación a las que tuvieron gestación simple (P < 0,05).
La producción de calostro también fue diferente entre los biotipos (P = 0,0023), ya que las M.M triplicaron la producción en relación a las C.C, siendo esta la mayor diferencia encontrada entre los diferentes biotipos. En la producción de calostro tampoco se encontraron diferencias en relación al tipo de parto coincidiendo con estudios realizados por Pattinson et al. (1995). En las ovejas F.(FxF.C) fue más frecuente encontrar calostro con mayor viscosidad, con tonos de amarillo más oscuro, mientras que M.M y C.C tuvieron calostros más líquidos con tonos más claros (P < 0,0001).
La composición del calostro fue similar para los distintos biotipos (grasa: 14,8 a 18,1 %; lactosa 3,1 a 3,6 %, proteína 18,7 - 21,9 % y sólidos no grasos 22,9 a 26,3 %). Sin embargo, la carga fetal afectó la composición (P = 0,0199), particularmente la concentración de proteína siendo 20,5 ± 0,7 % y 22,4 ± 1,1 % para ovejas que gestaron mellizos y trillizos vs. 18,1 ± 0,9 % para ovejas que gestaron únicos. Lo mismo sucedió para la concentración de sólidos no grasos (P = 0,0190; 24,8 ± 0,6 % y 26,3 ± 0,9 % para ovejas que gestaron mellizos y trillizos vs. 22,5 ± 0,8 % para ovejas que gestaron únicos). Estos datos se condicen a los observados por Banchero et al. (2010), quienes reportan calostros con 17,9 % y 20,7 % de proteína para ovejas con corderos únicos y mellizos respectivamente.
Sin embargo, Pattinson et al. (1995) observaron que la concentración de grasa, proteína total, IgG, cenizas y sólidos totales no eran afectados significativamente por el tamaño de la camada. Al calcular los gramos producidos de los principales componentes del calostro, las M.M producen más grasa, proteína, lactosa y sólidos no grasos en comparación con el resto de los biotipos (P = 0,0188, P = 0,0093, P = 0,0179 y P = 0,0086, respectivamente), explicado por el mayor volumen de calostro producido. Asimismo, el biotipo M.M produjo entre 64 y 75 % más sólidos totales que los biotipos M.C y M.(F.M). Por otro lado, la M.M produjo 142, 177 y 193 % más de sólidos totales que los biotipos F.C, C.C y F.(FxF.C).
Placenta: peso, cantidad y peso de los cotiledones y eficiencia placentaria
El tipo de parto y el biotipo afectaron el peso de la placenta (P = 0,0214 y P < 0,0001 respectivamente). Las ovejas de parto único presentaron placentas más livianas (0,43 ± 0,07 kg) seguidas por las melliceras y trilliceras (0,7 ± 0,04 kg y 1,0 ± 0,08 kg respectivamente). Esto estuvo directamente relacionado al PV total de la camada, representando la placenta entre el 8,8 y 9,2 % del peso de la misma. Dentro de los biotipos, las ovejas con sangre Milchschaf y las C.C fueron las que presentaron mayor peso de placenta (Cuadro 3). Esto seguramente también responda a la relación PV de los corderos con el peso de la placenta, donde se ha descrito una asociación entre el peso de la placenta y el peso al nacer del cordero alta, positiva y de tipo cuadrática (Montossi et al., 2005).
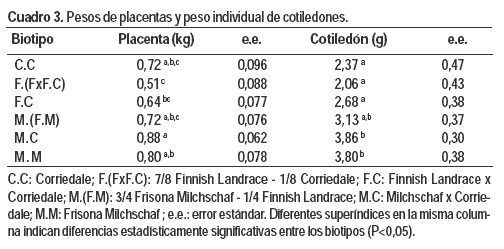
El número de cotiledones y el peso total de los mismos fue afectado sólo por el tipo de parto (P = 0,0087 y P = 0,0004 respectivamente). Las placentas de las ovejas con cordero único presentaron entre 25 y 30 % menos cotiledones (n = 66 ± 6) que las ovejas con corderos mellizos o trillizos, las cuales no difirieron entre sí. Esto obedece a que el número potencial de cotiledones es limitado porque el número de carúnculas en la oveja es finito (80 a 90; Botella, 1993), y el mayor número se alcanza con gestaciones gemelares sin observarse incremento con mayores cargas fetales (Dwyer et al., 2005). En cuanto al peso total de los cotiledones, las ovejas con tres corderos presentaron el mayor peso (0,33 ± 0,032 kg), siendo este 37 y 135 % superior al de las ovejas con corderos mellizos y únicos (P = 0,0024 y P = 0 ,0001 respectivamente).
El peso individual de los cotiledones estuvo afectado por el biotipo (P = 0,0033) y el tipo de parto (P = 0,0398). En general las placentas de biotipos con sangre Milchschaf tuvieron cotiledones más pesados. Por otro lado, las ovejas con tres corderos presentaron el mayor peso individual de cotiledones (3,78 ± 0,39 g.), siendo este 37 y 55 % superior al peso de los cotiledones de ovejas con corderos mellizos y únicos (P = 0,0237; P = 0,0161 respectivamente). No se encontraron diferencias significativas en el peso individual de los cotiledones de ovejas con cordero único y mellizo.
No se observaron diferencias estadísticamente significativas entre los diferentes biotipos y tipo de parto sobre la eficiencia placentaria y cotiledonaria. Esto obedece a lo explicado anteriormente, donde a menor número de cotiledones mayor peso de los mismos, dentro de cada biotipo.
Duración del parto, peso y comportamiento de los corderos en su primer hora de vida
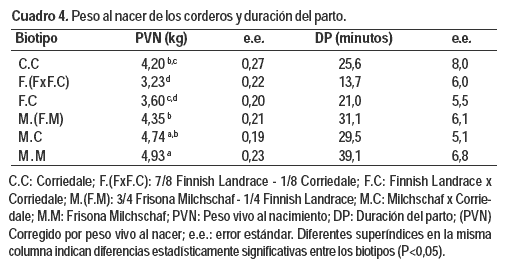
Con respecto a la DP, el tiempo que demoró en nacer cada cordero estuvo determinado por el tipo de nacimiento (P = 0,0194). Los corderos únicos demoraron 41,7 ± 6,9 minutos en nacer, (entre 22 y 23 minutos más que los nacidos de partos múltiples). Estas diferencias están explicadas por el PVN (P = 0,0088), por cada kilogramo que aumentó el PVN, la DP aumentó 7,26 ± 2,71 minutos. Los corderos nacidos como únicos pesaron más que los de partos múltiples (P = 0,0007; 4,88 ± 0,24 kg para corderos únicos vs. 4,03 ± 0,11 y 3,62 ± 0,17 kg para mellizos y trillizos que no presentaron diferencias estadísticamente significativas entre ellos). Ni el biotipo de la madre ni el sexo de los corderos presentaron diferencias significativas sobre la DP.
El PVN estuvo afectado por el biotipo de la madre (Cuadro 4; P < 0,0001), a medida que aumentó el porcentaje de sangre Milchschaf también aumentó el PVN; contrariamente, a medida que aumentó el porcentaje de sangre Finnish el PVN disminuyó.
Con respecto al comportamiento del cordero (Cuadros 5 y 6), ninguna de las características estudiadas (i.e. IP, LP, IM y LM) se vio afectada por el biotipo de la madre, tipo de nacimiento ni sexo del cordero. Sin embargo, el PVN influyó sobre los tiempos en que los corderos se pararon y lograron mamar (P = 0,0247 y P = 0,0101). Por cada kilogramo extra de PVN demoraron 2,86 ± 1,25 y 10,54 ± 4,00 minutos menos en LP y LM respectivamente.
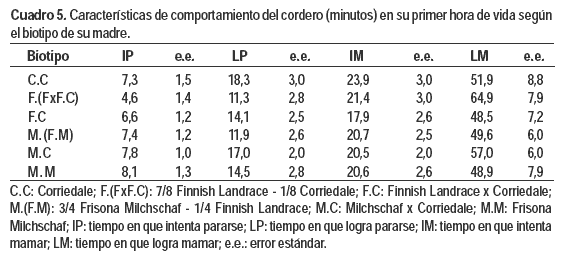
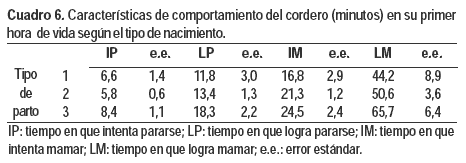
Conclusiones
El incremento de sangre Finnish en la cruza redundó en un aumento en la prolificidad con corderos más livianos y menor asistencia al parto. El uso de sangre Milchschaf en la cruza resultó en una prolificidad menor a la obtenida en cruzas con Finnish y una mayor asistencia al parto. La producción y composición de calostro fue similar en todos los biotipos cruza evaluados.
Consideramos que este trabajo hace un aporte al conocimiento del comportamiento de los nuevos biotipos ovinos prolíficos, manejados en condiciones nacionales, donde la información es escasa hasta el momento. Se requiere información objetiva del comportamiento de estos animales, tanto desde el punto de vista productivo como reproductivo, que permita generar tecnologías robustas. Estudios como este deberían repetirse con un número más alto de animales para poder contar con resultados que permitan extraer conclusiones con mayor confianza.
Bibliografía
Banchero G, Vázquez A, Quintans G, Ciappesoni G. 2014. Estudio preliminar del crecimiento, desarrollo e indicadores reproductivos de hembras de seis biotipos ovinos en Uruguay. Agrociencia (Uruguay), 18(2): 117 – 127.
Banchero G, Vázquez A, Montossi F, De Barbieri I, Quintans G. 2010. Pre-partum shearing of ewes under pastoral conditions improves the early vigour of both single and twin lambs. Animal Production Science, 50: 309 – 314.
Barbato G, Kremer R, Rosés L, Rista L. 2011. Producción de ovejas Corriedale y cruzas F1 con Milchschaf y Texel en condiciones de pastoreo. Veterinaria (Montevideo), 47(181): 9 – 13.
Bencini R, Purvis IW. 1990. The yield and composition of milk from Merino sheep. Proceedings of the Australian Society of Animal Production, 18: 144 – 147.
Bianchi G, Caravia V, Garibotto G, Bentancour O. 2000. Estudio comparativo de razas de lana blanca (Texel, Ile de France y Milchschaf) en la generación de madres cruza Corriedale [Cd-Rom]. En: XVI Reunión Latinoamericana de Producción Animal. III Congreso Uruguayo de Producción Animal; 28 - 31 de Marzo; Montevideo, Uruguay.
Botella J. 1993. La placenta : Fisiología y patología. Madrid : Ediciones Díaz de Santos. 390p.
Ciappesoni G, Vázquez A, Banchero G, Ganzábal A. 2014. Aumento de la prolificidad en la raza Corriedale : el caso uruguayo. En: 15° Congreso Mundial de Corriedale; 21 - 22 de Julio, 2014; Buenos Aires, Argentina. Buenos Aires : Asociación Argentina de Criadores de Corriedale. pp. 9 – 16.
Doney JM, Peart JN, Smith WF, Louda F. 1979. A consideration of the technique for estimation of milk yield by suckled sheep and a comparison of estimates obtained by two methods in relation to the effect of breed, level of production and stage of lactation. Journal of Agricultural Science, 92: 123 – 132.
Dwyer CM, Calvert SK, Farish M, Donbavand J, Pickup HE. 2005. Breed, litter and parity effects on placental weight and placentome number, and consequences for the neonatal behaviour of the lamb. Theriogenology, 63(4): 1092 – 1110.
Dwyer CM, Lawrence AB. 1999. Does the behaviour of the neonate influence the expression of maternal behaviour in sheep. Behaviour,136: 367 – 389.
Ganzábal A. 2013. Impacto productivo y económico del uso de materiales prolíficos en la producción de corderos. En: Seminario Técnico Producción Ovina de Calidad, INIA Treinta y Tres. Montevideo : INIA. pp. 135 – 141.
Ganzábal A. 2005. Análisis de registros reproductivos en ovejas Corriedale. En: Seminario de actualización técnica : Reproducción Ovina, recientes avances realizados por el INIA. Montevideo : INIA. (Serie de Actividades de Difusión ; 401). pp. 69 – 82.
Gigena F, Vázquez A. 2005. Suplementación estratégica en ovejas Corriedale durante los últimos días de gestación para aumentar la producción de calostro [Tesis de Grado]. Montevideo : Facultad de Agronomía. Universidad de la República. 88p.
Goot H, Eyal E, Foote WC, Matthews. 1984. The reproductive performance of Finn crosses in accelerated lambing programmes. En: The promotion of prolific strains of sheep by nutritional and managerial means. Bet Dagan, Israel : BARD. pp. 6 - 22.
Grunert E, Bove S, Stopiglia A. 1971. Fisiología de la gestación. En: Guía de Obstetricia Veterinaria fisiología de la gestación. Buenos Aires : Editorial Universitaria de Buenos Aires. pp. 1 – 12.
Kremer R, Barbato G, Rista L, Rosés L, Perdigón F. 2010. Reproduction rate, milk and wool production of Corriedale and East Friesian×Corriedale F1 ewes grazing on natural pastures. Small Ruminant Research, 90: 27 – 33.
Lira R. 2007. Producción ovina mejorada en la zona austral de Chile. Boletín, (164): 17p.
MAFF. 1975. Energy Allowances and Feeding Systems for Ruminants. Technical Bulletin (London), 33: 79p.
McCance I, Alexander G. 1959. The onset of lactation in the Merino ewe and its modification by nutritional factors. Australian Journal of Agricultural Research, 10: 699 - 719.
Montossi F, De Barbieri I, Ciappesoni G, Ganzábal A, Banchero G, Luzardo S, San Julián R. 2013. Intensification, diversification, and specialization to improve the competitiveness of sheep production systems under pastoral conditions: Uruguay’s case. Animal Frontiers, 3(3): 28 – 35.
Montossi F, De Barbieri I, Dighiero A, Martínez H, Nolla M, Luzardo S, Mederos A, San Julián R, Zamit W, Levratto J, Frugoni JC, Lima G, Costales J. 2005. La esquila preparto temprana : una nueva opción para la mejora reproductiva ovina. En: Seminario de actualización técnica : Reproducción Ovina, recientes avances realizados por el INIA. Montevideo : INIA. (Serie de Actividades de Difusión ; 401). pp. 85 – 104.
Morley FWH, White DA, Kennedy PA, Davis IF. 1978. Predicting ovulation rate from live weight in ewes. Agricultural Systems, 3: 27- 45.
Murphy PM. 1999. Maternal behaviour and rearing ability of Merino ewes can be improved by strategic feed supplementation during late pregnancy and selection for calm temperament [PhD thesis]. Perth : The University of Western Australia. 562p.
Pattinson SE, Davies DAR, Winter AC. 1995. Changes in the secretion rate and production of colostrum by ewes over the first 24 h post partum. Animal Science, 61(1): 63 – 68.
Scales GH, Burton RN, Moss RA. 1986. Lamb mortality, birthweight and nutrition in late pregnancy. New Zealand Journal of Agricultural Research, 29: 75 – 82.
SUL. 2006. Proyecto de transferencia integral : algunos resultados de su ejecución. Lana Noticias, (143): 48p.
Thomson BC, Muir PD, Smith NB. 2004. Litter size, lamb survival, birth and twelve week weight in lambs born to cross-bred ewes. Proceedings of the New Zealand Grassland Association, 66: 233-237.

