










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
Related links
Share

 Permalink
PermalinkAgrociencia (Uruguay)
Print version ISSN 1510-0839On-line version ISSN 2301-1548
Agrociencia Uruguay vol.20 no.1 Montevideo June 2016
Micorrizas en gramíneas nativas: efecto de la fertilización fosfatada a largo plazo
García Silvina1, Pezzani Fabiana1, Rodríguez Andrea2, del Pino Amabelia3
1Universidad de la República, Facultad de Agronomía, Ecología. Departamento de Sistemas Ambientales. Garzón 810, 12900 Montevideo, Uruguay. Correo electrónico: fabiana@fagro.edu.uy
2Universidad de la República, Facultad de Agronomía, Microbiología. Departamento de Biología Vegetal. Garzón 810, 12900 Montevideo, Uruguay.
3Universidad de la República, Facultad de Agronomía,Fertilidad. Departamento de Suelos y Aguas. Garzón 780, 12900 Montevideo, Uruguay.
Recibido: 20/8/15 Aceptado: 20/11/15
Resumen
En Uruguay el campo natural (CN) es el ecosistema dominante, relevante para la producción ganadera extensiva. Está constituido principalmente por gramíneas, especies que muestran alta micorrización. Los contenidos de fósforo (P) del CN son bajos, por lo que se llevan a cabo prácticas de fertilización fosfatada. El impacto de esta práctica sobre la comunidad vegetal difiere entre especies. El objetivo del trabajo fue estudiar el efecto del P sobre la colonización micorrícica en dos gramíneas nativas con respuestas diferenciales al agregado de P: Paspalum dilatatum y Coelorhachis selloana. Se planteó que las dos especies vegetales presentarían diferente micorrización en respuesta al P, lo que podría explicar en parte sus diferentes persistencias en suelos fertilizados. Se utilizó un experimento de larga duración instalado en INIA Palo a Pique, con tres tratamientos: CN sin fertilización y con fertilización fosfatada media y alta. Se evaluó la micorrización (total y por estructuras fúngicas) en P. dilatatum (mantuvo su frecuencia en parcelas fertilizadas con P) y en C. selloana (redujo su frecuencia). La fertilización fosfatada afectó negativamente la micorrización de ambas gramíneas y dicho efecto dependió de la estación del año. Dado que estos resultados fueron independientes de la respuesta de la especie en la comunidad vegetal, existirían otros factores involucrados en determinar el desempeño de estas gramíneas en situaciones de incremento de P. En ambientes con mayor contenido de P habría un control fitocéntrico que limitaría la micorrización considerando el costo energético que esta interacción tiene para las plantas.
Palabras clave: PASPALUM DILATATUM, COELORHACHIS SELLOANA, CAMPO NATURAL, FÓSFORO, COLONIZACIÓN MICORRÍCICA
Summary
Mycorrhizae in Native Grasses: Long-term Effects of Phosphorus Fertilization
Natural grasslands (NG) are the major Uruguayan ecosystem and are relevant for extensive livestock production. They consist mainly of grasses with high mycorrhizal colonization. NG soils have low phosphorus (P) content, so fertilization practices are carried out. The impact of this practice on the vegetal community differs between species. The aim of this work was to study the effect of P on mycorrhizal colonization in two native grasses with differential responses to the addition of P: Paspalum dilatatum and Coelorhachis selloana. It was suggested that the two species would present different mycorrhizal response to P, which could partly explain its persistence in fertilized soil. We used a long-term experiment from INIA Palo a Pique, with three treatments: NG without fertilization, and with high and medium levels of P fertilization. We evaluated mycorrhizal colonization (total and by fungal structures) in P. dilatatum (which maintained its frequency in plots fertilized with P) and in C. selloana (which reduced its frequency). P fertilization negatively affected mycorrhizal interaction with both grasses, and this effect was dependent on the season. Since these results were independent of the response of the species in the plant community, we postulated that there would be other factors involved in determining the performance of these grasses in situations of increasing P. In environments with higher content of P there would be a phytocentric control that would limit the mycorrhization considering the energetic cost that this interaction has for plants.
Keywords: PASPALUM DILATATUM, COELORHACHIS SELLOANA, NATURAL GRASSLAND, PHOSPHORUS, MYCORRHIZAL COLONIZATION
Introducción
Las micorrizas arbusculares son el tipo de micorrizas más comunes; están presentes en una amplia variedad de ambientes siendo particularmente de interés en ecosistemas agrarios ya que inciden en la productividad de las plantas (Johnson et al., 1997; Lekberg y Koide, 2005; Hoeksema et al., 2010; van der Heijden et al., 2015). Representan una interacción simbiótica entre hongos micorrizógenos arbusculares (HMA) del phylum Glomeromycota y las raíces de la mayoría de las plantas vasculares (Smith y Read, 2008). Entre las estructuras fúngicas que se desarrollan en las células corticales de las raíces de las plantas, los arbúsculos son particularmente importantes ya que es donde se produce el intercambio de sustancias y nutrientes entre los simbiontes. Otras estructuras que se desarrollan en las raíces son: hifas, ovillos y vesículas. Las hifas son estructuras tubulares que forman el micelio cenocítico y se desarrollan tanto dentro de las raíces como en el suelo. Los ovillos son hifas que penetran en las células corticales y adquieren una conformación torneada; podrían también participar en el intercambio metabólico aunque su función aún no se conoce cabalmente. Las vesículas son estructuras globosas que almacenan lípidos y representan reservas para los hongos. La fracción o el porcentaje de raíz que es colonizado por cada una de estas estructuras fúngicas (que se pueden observar y cuantificar luego de un procedimiento de tinción) es el método más usado para evaluar la presencia de micorrizas a nivel radicular (Smith y Read, 2008).
Se ha demostrado que esta interacción podría otorgar ciertos beneficios a las plantas hospedantes entre los que se destacan el aumento en la absorción de agua (Faber et al., 1991; Marulanda et al., 2003) y nutrientes, particularmente P (Sanders y Tinker, 1971; Jayachandran y Shetty, 2003; Smith y Smith, 2011) por tratarse este de un elemento inmóvil en el suelo. La planta por su parte destina entre el 10 y el 60 % de los carbohidratos que produce por fotosíntesis a mantener esta interacción (Stribley et al., 1980). Se ha encontrado que los beneficios de las micorrizas arbusculares para las plantas son más importantes en ambientes pobres en nutrientes así como con estrés hídrico (Bethlenfalvay et al., 1988; Tuomi et al., 2001; Pezzani et al., 2006; Martínez y Pugnaire, 2009). En este sentido, Bethlenfalvay et al. (1983) y Bethlenfalvay y Dakessian (1984) demostraron experimentalmente que el resultado de la interacción entre HMA y las plantas depende en gran medida de las concentraciones de P disponible en el suelo y los beneficios para las plantas sólo se observaron en un rango estrecho de concentración de P (4 -12 mg kg-1). Trabajos realizados en gramíneas y otras especies herbáceas demostraron que la fertilización fosfatada afectó negativamente la colonización micorrícica total así como las estructuras fúngicas, particularmente la presencia de arbúsculos (Covacevich et al., 1995; Aguilar et al., 2004; Chen et al., 2014).
En comunidades de pastizales se ha encontrado que las micorrizas arbusculares juegan un papel en la estructuración de las mismas, pudiendo tanto aumentar (Grime et al., 1987; van der Heijden et al., 1998, 2006; Karanika et al., 2008) como disminuir la diversidad de la comunidad vegetal (Hartnett y Wilson, 1999). En los pastizales las gramíneas constituyen la familia predominante y numerosos antecedentes han demostrado una importante presencia de micorrizas en estas especies (Hartnett y Wilson, 1999, 2002; Endresz et al., 2013).
En Uruguay el pastizal, conocido como campo natural, es el ecosistema dominante, ocupando aproximadamente el 68 % del territorio nacional (MGAP, 2011), siendo además el sustento de una de las principales actividades económicas del país como lo es la ganadería extensiva. En los suelos de campo natural tanto el N como el P son nutrientes limitantes para el crecimiento vegetal (Berretta et al., 1998). Particularmente, los contenidos de P que se han reportado son bajos, con valores que oscilan entre 3 y 5 mg kg-1 de suelo (Hernández et al., 1995). Con el fin de levantar estas restricciones, desde la década de 1950, se lleva a cabo una práctica agronómica conocida como «mejoramientos extensivos» que incluye la siembra de alguna especie de leguminosas y la fertilización fosfatada.
De acuerdo al último censo agropecuario estos mejoramientos representan aproximadamente el 5 % de la superficie de campo natural (MGAP, 2011). Si bien se reconoce el impacto positivo de esta práctica sobre la productividad forrajera a corto plazo, existen evidencias que señalan la escasa persistencia de las especies introducidas así como cambios en la composición florística del campo natural (Millot et al., 1987). Estos cambios fueron cuantificados en un experimento de larga duración instalado en 1996 en el departamento de Treinta y Tres (Uruguay) en el que se observaron respuestas diferenciales en la frecuencia de gramíneas nativas, así como un aumento de especies exóticas como Cynodon dactylon y Lolium multiflorum. Luego de 10 años de fertilización fosfatada, algunas de las gramíneas nativas desaparecieron del tapiz (por ejemplo Paspalum notatum, Stipa neesiana), otras mantuvieron su frecuencia (como Paspalum dilatatum y Panicum hians), mientras que otras disminuyeron su presencia en la comunidad sin llegar a desaparecer (por ejemplo Coelorhaquis selloana) (Jaurena et al., 2008a, 2008b). Considerando estas diferentes respuestas se planteó que las especies vegetales presentarían diferente micorrización en respuesta al P, lo que podría explicar en parte su persistencia en suelos fertilizados.
El objetivo general de este trabajo fue conocer el efecto de la fertilización fosfatada sobre la colonización por micorrizas arbusculares en dos gramíneas nativas del campo natural de Uruguay que presentaron respuestas diferenciales al agregado de P. En este contexto, se estudió cómo afecta el aumento en la disponibilidad de P la intensidad de colonización total y por estructuras de los HMA en P. dilatatum (especie considerada indiferente al P en términos de su frecuencia relativa) y en C. selloana (especie que decrece con P también considerando el cambio de su frecuencia). La hipótesis que se plantea es que en C. selloana la presencia de micorrizas se verá reducida a causa del incremento de P y que en P. dilatatum no será afectada.
Materiales y métodos
Sitio de estudio
El trabajo se realizó durante 2011 y 2012 en un experimento instalado en 1996 en la estación experimental Palo a Pique del Instituto Nacional de Investigaciones Agropecuarias (INIA) departamento de Treinta y Tres, Uruguay (33°14’58’’ S, 54°29’24’’ W). El suelo en el experimento corresponde a un Argisol Típico de la Unidad Alférez (Salvo et al., 2008).
Diseño experimental
El experimento consistió en tres tratamientos de fertilización fosfatada, con niveles medio y alto de P (PM y PA, respectivamente) y parcelas testigo de campo natural sin fertilizar (CN). Cuando se instaló el experimento, en las parcelas fertilizadas se sembró en cobertura Trifolium repens y Lotus corniculatus, especies que desaparecieron del tapiz vegetal luego de transcurridos 10 años (Jaurena et al., 2008a, 2008b).
Cada tratamiento tuvo cinco réplicas de 2 ha cada una, distribuidas en bloques al azar. Para el presente trabajo se seleccionaron tres bloques de acuerdo a la homogeneidad topográfica y edafológica, resultando en un total de nueve parcelas.
En el tratamiento con nivel medio de P (PM) se incorporaron 45 kg ha-1 de P2O5 a la siembra y refertilizaciones de 30 kg ha-1 de P2O5 y en el tratamiento con nivel alto de P (PA) se fertilizó con 90 kg ha-1 de P2O5 a la siembra y refertilizaciones de 60 kg ha-1 de P2O5. Las refertilizaciones se realizaron anualmente, en otoño. El Cuadro 1 muestra el contenido de P disponible (P Bray I) en mayo y noviembre de 2011 en las parcelas seleccionadas (Cuadro 1).

Muestreo de plantas
Los criterios con los cuales se seleccionaron las especies para el presente estudio fueron: a) haber tenido una respuesta diferencial de su frecuencia relativa frente a las situaciones de fertilización fosfatada; b) presentar alta frecuencia en las parcelas y buena calidad forrajera y c) estar presentes en todas las parcelas que se incluyeron en este trabajo. Con estos criterios se seleccionó a C. selloana, la cual redujo su frecuencia en parcelas fertilizadas (especie decreciente al P) y a P. dilatatum que no varió su frecuencia con el incremento de P (especie indiferente al P; Jaurena et al., 2008a, 2008b).
Se realizaron muestreos estacionales para evaluar la micorrización de las plantas en los meses de febrero (verano), mayo (otoño), agosto (invierno) y noviembre (primavera) de los años 2011 y 2012. En particular para los años 2011 y 2012 las precipitaciones acumuladas fueron de 1497 y 1045 mm respectivamente, mientras que las temperaturas promedio fueron de 16,6 °C en el 2011 y 17,3 °C en el año 2012 (INIA, 2015).
Previo a los muestreos se realizó un manejo del pastoreo de manera de homogeneizar la altura del tapiz vegetal en las nueve parcelas estudiadas. Para ello se establecieron cargas ganaderas altas y de poca duración de un rodeo vacuno de la Estación Palo a Pique. La altura remanente del tapiz fue de aproximadamente 15 cm al momento de hacer los muestreos. De cada una de las nueve parcelas y en cada fecha se extrajeron muestras de raíces de cinco individuos de P. dilatatum y de C. selloana seleccionados al azar considerando una distancia mínima entre ellos no menor a 5 m. Las raíces fueron conservadas en vinagre hasta su procesamiento en el laboratorio.
Colonización micorrícica
Las raíces colectadas fueron lavadas y aclaradas con una solución de KOH al 10 % y posteriormente teñidas con azul de tripano (0,05 %) según el método de Koske y Gemma (1989). Para cada planta se montaron en portaobjetos 30 segmentos de raíz de 1 cm de largo. Mediante observación al microscopio se identificaron las estructuras características de los HMA (arbúsculos, hifas, ovillos y vesículas). Para cada planta se cuantificó el porcentaje de colonización micorrícica total así como el porcentaje de las diferentes estructuras de los HMA a través del cociente de campos con presencia de alguna estructura fúngica y el número de campos observados.
Análisis estadístico
El modelo utilizado para el análisis de los resultados fue un Modelo Lineal Generalizado Mixto (MLGM), familia binomial, utilizando el programa estadístico InfoStat 2015. Se realizó un análisis de varianza de medidas repetidas con los factores: año, estación de muestreo y tratamiento para las variables de respuesta: colonización total; colonización por estructuras de intercambio (arbúsculos y ovillos) y colonización por vesículas. Las pruebas a posteriori de comparación de medias se realizaron mediante el test de Di Rienzo, Guzmán y Casanoves (DGC) y se trabajó con una significancia de 0,05.
Y= µ + Ai + Ej +Tk + (AxT)jk + (ExT)jk + (AxExT)ijk
i= 1,2 j=1,2,3,4 k=1,2 ,3
donde:
Y= variable de respuesta
µ=media general
Ai= efecto del año
Ej= efecto de la estación de muestreo
Tk= efecto del tratamiento
(AxT)ik= efecto de la interacción Año x Tratamiento
(ExT)jk= efecto de la interacción Estación x Tratamiento
(AxExT)ijk = efecto de la interacción Año x Estación x Tratamiento
Resultados
La fertilización fosfatada afectó negativamente la mayoría de las variables analizadas sobre la interacción micorrícica para las dos gramíneas estudiadas y dicho efecto fue dependiente en todos los casos de la estación de muestreo. El factor año mostró efectos significativos para la mayoría de las variables, a excepción de la colonización por arbúsculos. De acuerdo al modelo utilizado, no se observó un efecto significativo de la triple interacción año x estación x tratamiento para ninguna de las variables analizadas (Cuadro 2).
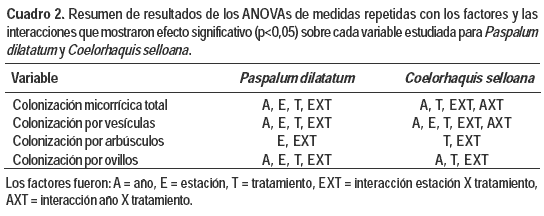
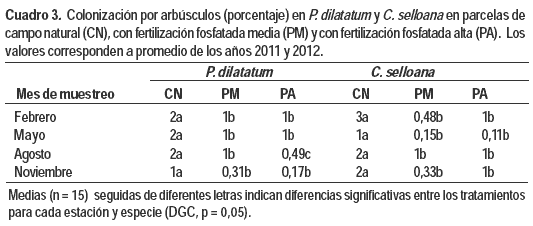
Si bien los niveles de P disponible en suelo son significativamente diferentes entre los tratamientos (Cuadro 1), dichas diferencias no se vieron reflejadas en la mayoría de las variables referidas a la micorrización entre las plantas provenientes de las parcelas con fertilización media y alta (Figura 1 y Cuadro 3).
En relación a la colonización micorrícica total, las plantas de P. dilatatum presentaron alta presencia de micorrizas en sus raíces en ambos años y en todas las estaciones. Para esta variable y esta especie se observó un efecto significativo del año (p<0,0001), de la estación (p<0,0001), del tratamiento (p<0,0001) y de la interacción estación x tratamiento (p= 0,0026) (Cuadro 2). Esta especie en 2011 mostró valores de colonización micorrícica total superiores a los de 2012, con un promedio de 88,5 y de 81,6 %, respectivamente (Figura 1a y 1c). En todas las estaciones la presencia de micorrizas fue significativamente mayor en las parcelas de CN en comparación con los tratamientos con fertilización. P. dilatatum mostró mayor presencia de micorrizas en febrero (verano) momento en el cual el efecto negativo del P fue el más bajo provocando una reducción del 2 % en el año 2011 y de 4,4 % en el 2012. La reducción de la colonización debida al P fue mayor en noviembre durante el 2011 y en agosto en 2012, donde se observaron decrementos del 21 y 25 % de la colonización micorrícica total respectivamente.
En C. selloana la colonización total también fue importante y difirió significativamente entre años (p<0,0001), teniendo un promedio de 84,8 % y 74,5 % en 2011 y 2012, respectivamente (Figura 1b y 1d). Se observó además un efecto significativo de la estación del año (p<0,0001), así como del tratamiento (p<0,0001), de la interacción estación x tratamiento (p<0,0001) y de la interacción año x tratamiento (p<0,0001) (Cuadro 2). También para esta especie, los mayores valores de la presencia de HMA se observaron en las parcelas de CN en todas las estaciones en 2011 y 2012. En agosto de 2011 y en febrero de 2012 C. selloana presentó la mayor presencia de micorrizas y el efecto negativo del P fue mayor en noviembre (primavera) en ambos años.
En ambas gramíneas se observó escasa presencia de arbúsculos en todas las muestras analizadas (registros menores al 5 % en todos los casos) (Cuadro 3). En P. dilatatum, la presencia de arbúsculos fue afectada significativamente por la estación (p<0,0001) y la interacción estación x tratamiento (p = 0,0063) (Cuadro 2). Para todas las épocas del año los registros de arbúsculos fueron superiores en las parcelas de CN que en las fertilizadas con valores máximos en febrero (verano). En agosto se observó la mayor reducción del nivel de arbúsculos debido al P con decrementos del 75 % (Cuadro 3). La colonización por arbúsculos en C. selloana fue afectada significativamente por el tratamiento (p<0,0001) y la interacción estación x tratamiento (p = 0,0086) (Cuadro 2). Los mayores niveles de estas estructuras, al igual que en P. dilatatum, se vieron en CN, con un registro máximo también en febrero (Cuadro 3).
La presencia de ovillos fue escasa en ambas gramíneas en todas las muestras analizadas, sin embargo se observaron valores superiores de estas estructuras fúngicas en relación a los arbúsculos. La colonización por ovillos fue afectada por casi todos los factores e interacciones (Cuadro 2). En general la adición de P provocó una disminución en la presencia de ovillos, aunque no existió un patrón tan definido como en el caso de los arbúsculos ya que hubo variaciones entre años y estaciones. En este sentido es destacable señalar que en el caso de P. dilatatum los mayores registros de ovillos ocurrieron en plantas sin fertilizar en febrero de 2011 (6 % ± 0,4) y en mayo de 2012 (5 % ± 0,2). Por su parte en C. selloana la mayor ocurrencia de ovillos se dio también en plantas de CN en febrero de ambos años (2011: 17 % ± 0,1; 2012: 5 % ± 0,1).
La presencia de vesículas de hongos micorrizógenos alcanzó valores elevados en ambas especies, entre 27 % y 68 %. Para P. dilatatum el máximo fue 48 % en agosto de 2011 y para C. selloana 68 % en febrero de 2011 respectivamente, aunque la variación en ambos casos fue muy importante debido a varios de los factores analizados. En P. dilatatum la colonización por vesículas fue afectada por el factor año (p<0,0001), estación (p<0,0001), tratamiento (p = 0,0012), y por la interacción entre tratamiento y estación (p = 0,0012). En ambos años y en la mayoría de las estaciones existió una mayor presencia de vesículas en CN en relación a las parcelas fertilizadas. En C. selloana, la presencia de vesículas fue afectada por el año (p<0,0001), el tratamiento (p = 0,0013) la interacción tratamiento x estación (p<0,0001) y la interacción año x tratamiento (p<0,0001). También para esta especie la mayor presencia de estas estructuras fúngicas se presentó en las parcelas de CN (excepto en agosto 2011 y mayo 2012).
Discusión
Los resultados obtenidos permitieron rechazar la hipótesis planteada dado que se constató un efecto negativo sobre la interacción micorrícica debido al incremento de P disponible en ambas gramíneas. De acuerdo a lo esperado, en C. selloana, la especie que redujo su frecuencia en respuesta al aumento en la disponibilidad de P, la interacción micorrícica fue negativamente afectada por la fertilización. La menor presencia de esta especie en el pastizal fertilizado podría explicarse entre otros factores por la reducción de la micorrización. En P. dilatatum, contrariamente a lo esperado, el P también afectó negativamente la micorrización. En esta especie hipotetizamos que el incremento de P no disminuiría la presencia de micorrizas, considerando que el resultado de esta interacción podría beneficiar a las plantas permitiendo así mantener la frecuencia de esta especie en la comunidad. Por lo tanto, en el caso de P. dilatatum existirían otros factores y procesos involucrados en determinar el desempeño de las plantas de esta especie en el pastizal en situaciones de incremento de P.
Dado que la interacción micorrícica arbuscular se trata de una simbiosis obligada para el hongo (el cual permanecería en forma de espora si no colonizara una planta hospedadora) se sugiere que la misma es comandada por la planta (Smith y Read, 2008; Smith y Smith, 2011). Anderson et al. (1994), Tuomi et al. (2001) y Chen et al. (2014) proponen que las plantas permitirían que la colonización se estableciera sólo en situaciones de escasez de nutrientes en las que las micorrizas aumentarían la capacidad de acceder a los mismos. Esto podría explicar la reducción de la colonización micorrícica en las situaciones de alta disponibilidad de P. Otra posible explicación de los resultados puede relacionarse con los cambios en la composición florística ocurridos en la comunidad vegetal del pastizal en el ensayo de larga duración en Palo a Pique. En las parcelas fertilizadas se registró un aumento de gramíneas invasoras como Lolium multiflorum y Cynodon dactylon, las cuales pueden haber alterado las comunidades de HMA de las gramíneas nativas, reduciendo la abundancia de dichos hongos como estrategia para establecerse en el nuevo ambiente (Wolfe y Klironomos, 2005; Hawkes et al., 2006; Pezzani et al., 2012; Endresz et al., 2013). En este sentido resulta interesante analizar a futuro si las gramíneas invasoras de esta comunidad presentan micorrizas y si dicha interacción es afectada por la fertilización fosfatada.
Nuestros resultados concuerdan con trabajos previos, realizados con gramíneas y especies herbáceas de otras familias, que reportaron efectos negativos del incremento de P sobre la micorrización (Covacevich et al., 1995; Aguilar et al., 2004; Chen et al., 2014).
Luego de 15 años de iniciado el experimento en el cual se desarrolló este estudio los valores de P disponible en suelo reflejaron los tratamientos de fertilización a los que fueron sometidas las parcelas. En este sentido se encontró que en el tratamiento de fertilización con niveles medios (PM) el contenido de P disponible aproximadamente triplicó el de las parcelas testigo sin fertilizar (CN) y que en el caso del tratamiento de alta dosis (PA) el P disponible en suelo fue seis y cuatro veces mayor en relación al testigo en mayo y noviembre de 2011 respectivamente. El aumento observado en el P disponible entre mayo y noviembre podría estar relacionado con procesos de mineralización de P orgánico en la primavera tardía, así como con fenómenos vinculados a las condiciones de óxido-reducción del suelo en las diferentes estaciones (Hernández y Meurer, 2000). En la zona este de Uruguay los suelos presentan una importante diferenciación, con un horizonte subsuperficial de textura pesada, lo que lleva a que cuando ocurren precipitaciones se produzcan períodos más o menos extendidos de condiciones reductoras. En esas condiciones el Fe+3 pasa a Fe+2, aumentando la solubilidad de los compuestos de Fe y P y por lo tanto la disponibilidad de P para las plantas. Al volver el suelo a las condiciones de oxidación este proceso se revierte. Estas fluctuaciones se han observado en suelos de la zona este de Uruguay, donde se ubica el experimento de Palo a Pique. En el presente trabajo no es posible afirmar que ocurriera este tipo de fenómeno, ya que no se realizaron muestreos sucesivos ni se midió el contenido de agua del suelo durante el período estudiado. No obstante, teniendo en cuenta estos antecedentes, una posible explicación del cambio en el nivel de P disponible se podría relacionar a este fenómeno (Ferrando et al., 2002).
Sin embargo, a pesar de estas notorias diferencias en los contenidos de P en el suelo, los efectos sobre la micorrización sólo se observaron al comparar plantas que no recibieron fertilización con aquellas fertilizadas, no registrándose diferencias significativas para la mayoría de las variables estudiadas entre los tratamientos de fertilización media y alta. Los niveles de P disponible alcanzados en las parcelas PA están por encima del límite de 12 mg kg-1 que algunos antecedentes (Bethlenfalvay et al., 1983; Bethlenfalvay y Dakessian, 1984) señalaron como la máxima concentración de P en la cual la interacción micorrícica otorgó beneficios para las plantas. Como fue previamente mencionado, en ambientes con mayor contenido de P podría haber un control fitocéntrico que limitara la presencia de los hongos considerando el costo energético que la interacción tiene para las plantas (Smith y Smith, 2011; Tuomi et al., 2001; Chen et al., 2014).
Independientemente del efecto de la fertilización fosfatada, tanto las raíces de P. dilatatum como las de C. selloana mostraron altos valores de colonización por micorrizas arbusculares. Este resultado concuerda con estudios previos con estas mismas especies (Parodi y Pezzani, 2011; García, 2012) o con otras gramíneas que también son estivales y poseen metabolismo fotosintético C4 (Wilson y Hartnett, 1998; Lugo et al., 2003). En dichos trabajos se sugiere que la mayor micorrización de gramíneas C4 en relación a las de tipo C3 podría deberse a que las primeras presentan raíces más gruesas y las hifas del micelio desarrollado en la colonización por los HMA podrían comportarse como las raíces finas participando activamente en la absorción de agua y nutrientes.
Los resultados evidenciaron efecto del año para la mayoría de las variables analizadas. En ambas especies de gramíneas observamos una mayor presencia de micorrizas durante el año 2011 con respecto al 2012. Esto podría relacionarse con los mayores niveles de precipitaciones que en el 2011 fueron 30 % superiores a los ocurridos en 2012 (INIA, 2015). Esta mayor disponibilidad hídrica podría haber permitido un mayor crecimiento de las plantas, lo que redundaría en mayor energía disponible para ser destinada a los HMA. Sin embargo, existen múltiples procesos y factores que pueden verse afectados con el aumento en la disponibilidad hídrica.
Los resultados obtenidos demostraron una marcada estacionalidad en la presencia de micorrizas en ambas especies, lo que concuerda con los resultados obtenidos por Parodi y Pezzani (2011) en un trabajo realizado también con gramíneas nativas de Uruguay. Los mayores valores de colonización micorrícica total se detectaron en febrero (verano) tanto de 2011 como de 2012. Esto podría relacionarse con el ciclo fenológico de las especies estudiadas ya que al tratarse de gramíneas estivales en verano presentan el mayor desarrollo del área foliar (Millot et al., 1987; Carámbula, 2004), lo que les permitiría una mayor producción de fotoasimilados, parte de los cuales podrían asignarse a la interacción micorrícica. Otra evidencia del efecto estacional sobre las micorrizas lo constituye la mayor presencia, también en verano, de las estructuras fúngicas de intercambio: los mayores registros tanto de arbúsculos como de ovillos ocurrieron en febrero en ambas especies (en P. dilatatum también se observó una importante cantidad de ovillos en mayo 2012). Tanto los arbúsculos como los ovillos son estructuras de los hongos que están presentes durante pocos días en las células de la raíz (Toth, 1992). La mayor ocurrencia en época estival de estas estructuras también estaría apoyando la relación entre el funcionamiento de las micorrizas y el ciclo fenológico de las gramíneas estudiadas.
El efecto del nivel de P sobre las micorrizas varió a lo largo del año ya que para todas las variables analizadas la interacción estación x tratamiento fue significativa en ambas especies. Se observó que el efecto negativo del P fue menor en verano que en los restantes momentos del año. En noviembre (primavera) en ambas especies la micorrización presentó los valores más bajos y además fue la época del año en la que el aumento de P causó la mayor reducción de la presencia de los HMA. En las especies estivales desde finales de la primavera las plantas priorizan la asignación de energía a la función reproductiva (Millot et al., 1987; Carámbula, 2004) lo cual podría comprometer los recursos destinados al mantenimiento de la micorrización. A su vez los contenidos de P disponible en suelo fueron superiores en noviembre en relación a mayo (al menos para el 2011). Por lo tanto, la reducción en la micorrización de estas especies en primavera podría explicarse tanto por aspectos relacionados a la fenología de las plantas como por una mayor cantidad de P disponible en el ambiente.
Las plantas provenientes de las parcelas de campo natural presentaron más vesículas de hongos micorrizógenos que las plantas de parcelas fertilizadas por lo cual el incremento de P en suelo también afectó negativamente esta variable. Los resultados obtenidos coinciden con los de Smith y Read (2008) quienes reportaron efectos negativos sobre la presencia de vesículas en condiciones ambientales de mayor concentración de P y de baja radiación. Tanto en P. dilatatum como en C. selloana observamos niveles importantes de colonización por estas estructuras, superiores a 50 % en 2011, el año de mayores precipitaciones. No se encontraron antecedentes que analizaran la relación entre la disponibilidad hídrica y la presencia de estas estructuras.
Este trabajo representa la primera contribución en el estudio de la fertilización fosfatada a largo plazo sobre la micorrización en gramíneas nativas del campo natural de Uruguay. Plantea interrogantes que podrían abordar futuras investigaciones sobre el impacto que podría tener la reducción de la micorrización en el contenido de P en los tejidos vegetales así como en el crecimiento de las plantas y su desempeño en la comunidad vegetal.
Agradecimientos
Las autoras agradecen al Instituto Nacional de Investigación Agropecuaria (INIA) Treinta y Tres por el aval y apoyo logístico para la realización de este trabajo. Un particular agradecimiento a Graciela Quintans y al personal de la Estación Experimental Palo a Pique por permitirnos utilizar la infraestructura de la Estación y coordinar el pastoreo del ensayo. Agradecemos también a Felipe Lezama y Gerardo Parodi por la colaboración en el trabajo de campo y a Alejandra Borges por el asesoramiento en el análisis estadístico. Este trabajo fue financiado por el Proyecto 1814 del Fondo Clemente Estable – ANII.
Bibliografía
Aguilar C, Casanova O, Bordoli JM. 2004. Evaluación de la micorrización y la fertilización fosfatada en Lotus Rincón. En: Memorias de la XX Reunión del grupo Técnico Regional del Cono Sur en el Mejoramiento y Utilización de los Recursos Forrajeros del Área Subtropical y Tropical – Grupo Campos; Salto, Uruguay. Salto : UR. pp. 281 – 282.
Anderson RC, Hetrick BAD, Wilson GWT. 1994. Mycorrhizal dependence of Andropogon gerardii and Schizachyrium scoparium in two prairie soils. American Midland Naturalist, 132: 366 – 376.
Berretta EJ, Risso DF, Levratto JC, Zamit WS. 1998 Mejoramiento de campo natural de basalto fertilizado con nitrógeno y fósforo. En: Seminario de Actualización en tecnologías para Basalto. Montevideo : INIA. (Serie Técnica ; 102). pp. 63 – 73.
Bethlenfalvay G, Thomas RS, Dakkessian S, Brown MS, Ames RN. 1988. Mycorrhizae in stressed environments : effects on plant growth, endophyte development, soil stability and soil water. En: Whitehead E, Hutchinson C, Timmermann B, Varady RG. [Eds.]. Arid Lands : Today and Tomorrow. Boulder : Westview Press. pp. 1015 - 1035.
Bethlenfalvay GJ, Dakessian S. 1984. Grazing effects on mycorrhizal colonization and floristic composition of the vegetation on a semiarid range in northern Nevada. Journal of Range Management, 37: 312 – 316.
Bethlenfalvay GJ, Bayne HG, Pacovsky RS. 1983. Parasitic and mutualistic associations between mycorrhizal fungus and soybean : the effect of phosphorous on host plant-endophyte interactions. Physiologia Plantarum, 57: 543 – 548.
Carámbula M. 2004. Pasturas y forrajes, manejo, persistencia y renovación de pasturas. Montevideo : Hemisferio Sur. 413p.
Chen Y-L, Zhang X, Ye J-S, Han H-Y, Wan S-Q, Chen B-D. 2014 Six-year fertilization modifies the biodiversity of arbuscular mycorrhizal fungi in a temperate steppe in Inner Mongolia. Soil Biology and Biochemistry, 69: 371 – 381.
Covacevich F, Echeverria HE, Andreoli E. 1995. Micorrización vesículo-arbuscular espontánea en trigo en función de la disponibilidad de fósforo. Ciencia del Suelo, 13: 47 – 51.
Endresz G, Somodi I, Kalapos T. 2013. Arbuscular mycorrhizal colonization of roots of grass species differing in invasiveness. Community Ecology, 14: 67 – 76.
Faber BA, Zasoski RJ, Munns DN, Shackel K. 1991. A method for measuring hyphal nutrient and water uptake in mycorrhizal plants. Canadian Journal of Botany, 69: 87 – 94.
Ferrando M, Mercado G, Hernández J. 2002. Dinámica del hierro y disponibilidad de fósforo durante períodos cortos de anaerobiosis en los suelos. Agrociencia, 6: 1 – 9.
García S. 2012. Respuestas de la colonización micorrícica a los componentes del pastoreo en Paspalum dilatatum (Poir) [Tesis de grado]. Montevideo : Universidad de la República. 29p.
Grime JP, JM Mackey, SH Hillier, DJ Read. 1987. Floristic diversity in a model system using experimental microcosms. Nature, 328: 420 – 422.
Hartnett DC, Wilson GWT. 2002. The role of mycorrhizas in plant community structure and dynamics : lessons from grasslands. Plant and Soil, 244: 319 – 331.
Hartnett D, Wilson T. 1999. Mycorrhizae influence plant community structure and diversity in tallgrass prairie. Ecology, 80: 1187 – 1195.
Hawkes C V, Belnap J, D’Antonio C, Firestone MK. 2006. Arbuscular mycorrhizal assemblages in native plant roots change in the presence of invasive exotic grasses. Plant and Soil, 281: 369 – 380.
Hernandez J, Meurer EJ. 2000. Phosphorus availability in six Uruguayan soils affected by alternation of the flooded-drained conditions. Revista Brasileira de Ciência do Solo, 24: 19 – 26.
Hernández J, Otegui O, Zamalvide JP. 1995. Formas y contenidos de fósforo en algunos suelos de Uruguay. Boletín de Investigaciones (Facultad de Agronomía), (43): 31p.
Hoeksema JD, Chaudhary VB, Gehring CA, Johnson NC, Karst J, Koide RT, Pringle A, Zabinski C, Bever JD, Moore JC. 2010. A meta-analysis of context-dependency in plant response to inoculation with mycorrhizal fungi. Ecology Letters, 13: 394 – 407.
INIA. 2015. Banco de Información Agroclimática [En línea]. Consultado 23 febrero 2015. Disponible en: http://www.inia.uy/investigaci%C3%B3n-e-innovaci%C3%B3n/unidades/GRAS/Clima/Banco-datos-agroclimatico.
Jaurena M, Ayala W, Terra J, Bermúdez R, Lagomarcino X. 2008a. Impacts of long-term phosphorus fertilization and addition of perennial legumes on a temperate natural grassland : I. Changes in species biodiversity and stability. En: XXI International Grassland and VIII International Rangeland Congress; 28 - 29 junio, 2008; Hohot, Inner Mongolia, China. Vol. 1. Hohot : Guangdong People’s Publishing House. pp. 82.
Jaurena M, Salvo L, Terra J, Ayala W, Bermúdez R, Barrios E, Lezama F. 2008b. Cambios en el largo plazo de características de suelo y vegetación de mejoramientos de campo natural. En: XXII Reunión del Grupo Técnico en Forrajeras del Cono Sur (Grupo Campos); 21 - 23 octubre, 2008; Minas, Uruguay. Montevideo : INIA. pp. 188.
Jayachandran K, Shetty KG. 2003. Growth response and phosphorus uptake by arbuscular mycorrhizae of wet prairie sawgrass. Aquatic Botany, 76: 281 – 290.
Johnson NC, Graham JH, Smith FA. 1997. Functioning of mycorrhizal associations along the mutualism-parasitism continuum. New Phytologist, 135: 575 – 585.
Karanika ED, Mamolos AP, Alifragis DS, Kalburtji KL, Veresoglou DS. 2008. Arbuscular mycorrhizas contribution to nutrition, productivity, structure and diversity of plant community in mountainous herbaceous grassland of northern Greece. Plant Ecology, 199: 225 – 234.
Koske R, Gemma J. 1989. A modified procedure for staining roots to detect VA mycorrhizas. Mycological Research, 92: 486 – 505.
Lekberg Y, Koide RT. 2005. Is plant performance limited by abundance of arbuscular mycorrhizal fungi? A meta-analysis of studies published between 1988 and 2003. New Phytologist, 168: 189 – 204.
Lugo M, González Maza M, Cabello M. 2003. Arbuscular mycorrhizal fungi in a mountain grassland II : seasonal variation of colonization studied, along with its relation to grazing and metabolic host type. Mycologia, 93: 407 – 415.
Martínez L, Pugnaire F. 2009. Interacciones entre las comunidades de hongos formadores de micorrizas arbusculares y de plantas. Algunos ejemplos en los ecosistemas semiáridos. Revista Ecosistemas, 18: 44 – 54.
Marulanda A, Azcón R, Ruiz-Lozano JM. 2003. Contribution of six arbuscular mycorrhizal fungal isolates to water uptake by Lactuca sativa plants under drought stress. Physiologia Plantarum, 119: 526 – 533.
MGAP. 2011. Censo General Agropecuario 2011 : resultados definitivos [En línea]. Consultado 16 julio 2015. Disponible en: http://www.mgap.gub.uy/portal/page.aspx?2,diea,diea-censo-2011-resultados-definitivos.
Millot JC, Risso D, Methol R. 1987. Relevamiento de pasturas naturales y mejoramientos extensivos en áreas ganaderas del Uruguay : Informe técnico para la Comisión Honoraria del Plan Agropecuario. Montevideo : MAP. 199p.
Parodi G, Pezzani F. 2011. Micorrizas arbusculares en dos gramíneas nativas de Uruguay en áreas con y sin pastoreo. Agrociencia, 15: 1 – 10.
Pezzani C, del Pino A, Lezama F, Rodriguez M, Garcia S, Parodi G. 2012. Interacciones complejas : micorrizas arbusculares y fósforo en pastizales naturales de Uruguay [En línea]. En: VII Simposio Nacional y IV Reunión Iberoamericana de la Simbiosis Micorrícica; 27 - 30 mayo, 2012; Xalapa. México. Consultado 19 febrero 2016. Disponible en: http://www.uv.mx/Simbiosis-Micorrizica/resumenes.html.
Pezzani F, Montaña C, Guevara R. 2006. Associations between arbuscular mycorrhizal fungi and grasses in the successional context of a two-phase mosaic in the Chihuahuan Desert. Mycorrhiza, 16: 285-295.
Salvo L, Terra J, Ayala W, Bermudez R, Correa J, Avila P, Hernández J. 2008. Long-term phosphorous fertilization and perennials legumens addition impacts on a températe natural grassland : II. Total and particulate soil organic carbon. En: XXI International Grassland and VIII International Rangeland Congress; 28 - 29 junio 2008; Hohot, Inner Mongolia, China. Vol. 1. Hohot : Guangdong People’s Publishing House. pp. 382.
Sanders FE, Tinker PB. 1971. Mechanism of absorption of phosphate from soil by Endogone mycorrhizas. Nature, 233: 278 – 279.
Smith SE, Smith FA. 2011. Roles of arbuscular mycorrhizas in plant nutrition and growth : new paradigms from cellular to ecosystem scales. Annual Review of Plant Biology, 62: 227- 250.
Smith SE, Read DJ. 2008. Mycorrhizal Simbiosis. 3rd ed. New York : Academic Press. 787p.
Stribley DP, Tinker PB, Rayner JH. 1980. Relation of internal phosphorus concentration and plant weight in plants infected by vesicular arbuscular mycorrhizas. New Phytologist, 86: 261 – 266.
Toth R. 1992. The quantification of arbuscules and related structures using morphometric cytology. En: JR Norris, DJ Read, Varma AK. [Eds.]. Methods in Microbiology. Vol. 24. London : Academic Press. pp. 275 – 299.
Tuomi J, Kytöviita MM, Härdling R. 2001.Cost efficiency of nutrient acquisition and the advantage of mycorrhizal symbiosis for the host plant. Oikos, 92: 62 – 70.
van der Heijden M, Martin F, Selosse MSI, Sanders I. 2015. Mycorrhizal ecology and evolution : the past, the present, and the future. New Phytologist, 205: 1406–1423.
van der Heijden M, Streitwolf-Engel R, Riedl R, Siegrist S, Neudecker A, Ineichen K, Boller T, Wiemken A, Sanders I.2006. The mycorrhizal contribution to plant productivity, plant nutrition and soil structure in experimental grassland. New Phytologist, 172: 739 – 752.
van der Heijden M, Boller T, Wiemken A, Sanders IR. 1998. Different arbuscular mycorrhizal fungal species are potential determinants of plant community structure. Ecology, 79: 2082 – 2091.
Wilson G, Hartnett D. 1998. Interespecific variation in plant responses to mycorrhizal colonization in tallgrass praire. American Journal of Botany, 85: 1732 – 1738.
Wolfe BE, Klironomos JN. 2005. Breaking new ground: soil communities and exotic plant invasion. Bioscience, 55: 477 – 487.

