










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Links relacionados
Compartilhar

 Permalink
PermalinkAgrociencia (Uruguay)
versão impressa ISSN 1510-0839versão On-line ISSN 2301-1548
Agrociencia Uruguay vol.19 no.2 Montevideo dez. 2015
Efecto de la fertilización fosfatada sobre la composición química de Lotus tenuis y Trifolium repens
Silveira David1, Hernández Jorge2, Cadenazzi Mónica3, del Pino Amabelia2, Zanoniani Ramiro4, Boggiano Pablo4
1Facultad de Agronomía, Universidad de la República, Departamento de Producción Animal y Pasturas. Estación Experimental Profesor Bernardo Rosengurtt, Ruta 26, km 408, 37000 Cerro Largo, Uruguay
Correo electrónico: edsilveira@fagro.edu.uy
2 Facultad de Agronomía, Universidad de la República, Departamento de Producción Animal y Pasturas. Estación Experimental Mario A. Cassinoni, Ruta 3, km 363, 60000 Paysandú, Uruguay
3 Facultad de Agronomía, Universidad de la República, Departamento de Suelos y Aguas. Avenida Eugenio Garzón 780, 12900. Montevideo, Uruguay
4 Facultad de Agronomía, Universidad de la República, Departamento de Biometría Estadística y Cómputo. Estación Experimental Mario A. Cassinoni, Ruta 3, km 363, 60000 Paysandú, Uruguay
Recibido: 24/2/14 Aceptado: 31/7/15
Resumen
Una manera de aumentar el rendimiento y calidad de forraje de las pasturas nativas es a través de la inclusión de leguminosas en el tapiz. Para un establecimiento apropiado de las mismas es necesario contar con una disponibilidad adecuada de fósforo (P) en el suelo. Dados los niveles deficientes de este nutriente en muchos suelos, es necesario su aporte a través de la fertilización. El objetivo del presente estudio fue evaluar el efecto del P aplicado a la siembra en varias dosis sobre el contenido de P y nitrógeno (N) de Lotus tenuis Waldst. & Kit ex. Willd. (lotus de los bajos) y Trifolium repens L. (trébol blanco), en un Gleysol Lúvico Melánico Típico del Departamento de Cerro Largo, Uruguay. Se evaluaron cuatro niveles de P (0, 21,8, 43,6 y 65,4 kg de P.ha-1) aplicados al voleo en la siembra, como superfosfato triple (0-46-0). El diseño experimental fue de bloques al azar, con ocho repeticiones. A los 120 días post-siembra se recolectaron 10 plantas/parcela/especie, para determinar el peso seco de las partes aéreas y raíz y su contenido de P y N. La concentración de P en la materia seca aumentó con la dosis de P para ambas especies, en tanto que el contenido de N mostró diferencias significativas solo en T. repens.
Palabras clave: FERTILIZACIÓN FOSFATADA, RELACIÓN NITRÓGENO-FÓSFORO, ANÁLISIS DE PLANTAS, LOTUS TENUIS, TRIFOLIUM REPENS
Summary
Effect of Phosphorus Fertilization on the Chemical Composition of Lotus tenuis and Trifolium repens
One way of increasing forage yield and quality of natural pastures is through the inclusion of legumes in the grass cover. In order to achieve an appropriate crop establishment it is necessary to have an adequate availability of phosphorus (P) in the soil. Given the poor levels of this nutrient in many soils, it is necessary to fertilize. The objective of this study was to evaluate the effect of P fertilization at various doses on the content of P and nitrogen (N) in Lotus tenuis Waldst. & Kit ex. Willd. (lotus from the lowlands) and Trifolium repens L. (white clover) in a smectitic, fine, thermic Oxiaquic vertic argiudoll at Cerro Largo, Uruguay. The treatments were four phosphorus levels (0, 21,8, 43,6 and 65,4 kg P ha-1) broadcasted at seeding as triple superphosphate (0-46-0). The experimental design was randomized blocks, with eight repetitions. At 120 days post seeding, 10 plants/plot/specie were collected to determine the aerial part and root dry weight, and P and N content. P concentration in dry matter increased with P doses for both species, while N concentration showed significant differences only in T. repens.
Keywords: PHOSPHATE FERTILIZATION, NITROGEN-PHOSPHORUS RATIO, PLANT ANALYSIS, LOTUS TENUIS, TRIFOLIUM REPENS
Introducción
En los campos del Uruguay predominan las gramíneas estivales sobre las invernales, generando una distribución primavero-estivo-otoñal de la producción de forraje. Asimismo, las pasturas naturales presentan una ausencia parcial de especies de leguminosas (Carámbula, 1996, 1977; Millot et al., 1987). En los sistemas pastoriles que tienen como base forrajera el campo natural, la nutrición del ganado depende de la disponibilidad y composición química de esas pasturas, lo cual es función de la estación del año y condiciones climáticas de la misma, así como de las cargas ganaderas manejadas (Millot et al., 1987).
El principal factor que afecta el establecimiento y la sobrevivencia de las especies de leguminosas es la disponibilidad de fósforo (P) en el suelo. Los niveles de P en el suelo y la magnitud de la respuesta a la fertilización fosfatada determinan los niveles de fertilizante a agregar (Ayala Torales, 2000; Bordoli, 1998; Wedderburn et al., 1996). En Uruguay, los suelos son naturalmente deficientes en P, siendo sus niveles muy bajos para el desarrollo de leguminosas forrajeras (Hernández, 1999). Esto, asociado a una escasa o nula aplicación de P a pasturas del campo natural, hace que sea el principal factor limitante para la instalación de leguminosas sobre el tapiz o sembradas en mezclas con gramíneas
Para algunos nutrientes minerales, hay diferencias en la concentración entre la parte aérea y la raíz de las plantas forrajeras. Las diferencias dependen en parte de las funciones del elemento, y en parte de si el elemento es susceptible a ser limitado por el tejido de la raíz (Bermúdez et al., 2000).
En las leguminosas, a veces existe una concentración más alta de nitrógeno en la raíz (4,5-9 % N) que en la parte aérea, debido a la concentración elevada de este nutriente en los nódulos de la raíz a causa de una activa fijación de nitrógeno atmosférico. Dicha tendencia se observa también para el P y el azufre. En T. repens, la concentración crítica de nitrógeno (N) en hoja + pecíolo después de cuatro semanas pos pastoreo es aproximadamente 3,5 % y en las plantas enteras (incluidas las raíces) 15 semanas pos siembra es aproximadamente 3,2 % (Whitehead, 2000).
Las concentraciones críticas para el fósforo en varias gramíneas y especies de leguminosas han sido reportadas por varios autores. Las necesidades de P de las pasturas con leguminosas son generalmente mayores que las de los cultivos, debido a que los contenidos en la materia seca del forraje con un suministro adecuado de P varían entre un 0,22 y 0,45 % (Barbazán et al., 2007; Whitehead, 2000; Bordoli, 1998; Scott, 1981). La concentración crítica para P disminuye con la madurez de las plantas. Las estimaciones de concentración crítica de P para la parte aérea de las especies de Lotus spp varían. Para Lotus corniculatus la concentración de P oscila entre 2,3 y 3,5 g kg-1 materia seca (Frame et al., 1998).
En T. repens creciendo a campo las concentraciones de P son más altas en las raíces que en la parte aérea, sobre todo cuando a las plantas se les proporciona una dosis alta de fertilizante fosfatado. Un factor que contribuye a la variación en la proporción en el contenido de P en la raíz/parte aérea en las leguminosas puede ser el número de nódulos que estén presentes en la raíz (Whitehead, 2000; Hart, 1989).
El manejo de los niveles de nitrógeno y fósforo en leguminosas sembradas en cobertura en sistemas ganaderos extensivos con fertilizaciones fosfatadas constituye una alternativa tecnológica para aumentar la producción y calidad del forraje ofrecido (Carámbula, 2002; Bordoli, 1998; Carámbula, 1996). El objetivo planteado en este trabajo fue investigar el efecto de dosis crecientes de fósforo aplicadas a la siembra sobre la concentración de N y P y sus relaciones en plantas de Lotus tenuis y Trifolium repens.
Materiales y métodos
El experimento en condiciones de campo se localizó en la Estación Experimental Profesor Bernardo Rosengurtt (EEBR), Facultad de Agronomía, Universidad de la República, Cerro Largo, Uruguay (Latitud 32° 25’ S y Longitud 54° 15’ W, Altitud 177 metros sobre el nivel del mar). El suelo se clasificó como Gleysol Lúvico Melánico Típico (smectitic, fine, thermic Oxiaquic vertic argiudoll) de la Unidad de Suelos Palleros (DSF, 1976). El clima se caracteriza como templado cálido (subtropical húmedo), denominado Cf (Cfa); sistema de clasificación de clima de Köppen - Geiger (Kottek et al., 2006).
El experimento se desarrolló entre los meses de junio y octubre del 2003. Previo a la siembra cada una de las especies de leguminosas fue inoculada individualmente con su respectiva cepa de rhizobium (adherente + inoculante + semilla), y peleteada con hiperfosfato fino. El 9 de junio se realizó la siembra al voleo en cobertura en una densidad por parcela de 465 semillas viables por m2 de Lotus tenuis Waldst. et Kit. cv Larrañaga y de 1373 semillas viables por m2 de Trifolium repens L. cv Zapicán. Al momento de la siembra fueron aplicados los tratamientos que correspondieron a un testigo sin fertilizar, y tres dosis de fósforo equivalentes a 21,8, 43,6 y 65,4 kg de P ha-1. Las dosis de P fueron aplicadas al voleo como superfosfato triple de calcio (sólido granulado, Ca(H2PO4)2, grado equivalente 0-46-0; 13-15 % Ca). El diseño experimental fue en bloques completamente aleatorizados con parcelas de 165 m2 y 8 repeticiones.
Se tomaron 15 muestras de suelo por bloque, a la profundidad de 15 cm, para determinar el contenido de P nativo. Las muestras fueron secadas en estufa de aire forzado a 38 ºC por 48 horas y posteriormente se molieron y tamizaron a un tamaño menor de 2 milímetros. El P asimilable se determinó colorimétricamente mediante el método Bray I (Bray y Kurtz, 1945), siendo su valor promedio para el área del experimento de 8 mg kg-1.
Para estudiar el efecto de la dosis de P sobre el contenido de N y P en la materia seca y la relación N/P de las plantas, a los 120 días de la siembra se recolectaron diez plántulas por especie y por parcela. Cada cilindro de suelo con su respectiva plántula se la dejo en recipientes individuales con agua durante 6 horas, posteriormente se removió la tierra y las plántulas fueron lavadas en un recipiente con malla para evitar la pérdida de raíces y nódulos. Cada plántula se fraccionó en parte aérea y raíz, y ambas fracciones fueron secadas durante 48 horas a 60 ºC en una estufa de aire forzado y finalmente pesadas para establecer su peso seco. A continuación fueron molidas con un molino Micro-Waley y tamizadas a un tamaño menor a 0,5 milímetros. Las determinaciones de los niveles de N y P en planta consistieron en una digestión por vía húmeda de 0,5 g de material vegetal con ácido sulfúrico a 350 °C durante 60 minutos y posterior oxidación con perhidrol. La determinación del nitrógeno se realizó por el método de Kjeldahl y el fósforo fue determinado por colorimetría mediante el método de Murphy y Riley (1962). Los contenidos de N y P se expresaron como porcentaje de la materia seca.
A partir de estos valores y el correspondiente a la materia seca de cada fracción se determinó el contenido de N y P en la parte aérea (NPA y PPA respectivamente) y raíces (NR y PR respectivamente) para ambas especies, estableciéndose la relación P/N en plantas a los 120 días pos-siembra (dps).
El efecto de los tratamientos se detectó mediante la técnica de análisis de varianza, ajustándose regresiones lineales y polinomios segmentados cuadrático-plateau entre las variables respuesta y los niveles de fertilización fosfatada. La herramienta informática utilizada en el análisis estadístico fue S.A.S. v. 8.1 (S.A.S. Institute. Cary, USA).
Resultados y discusión
Lotus tenuis Waldst. et Kit.
a) Concentración de P de la planta entera (PPLE), parte aérea (PPA) y raíz (PR)
La concentración de PPLE en L. tenuis presentó una respuesta positiva y significativa con el incremento en las dosis de P agregado a la siembra (Figura 1). El aumento del PPLE fue resultado del aumento positivo y significativo que mostraron sus componentes PPA y PR.
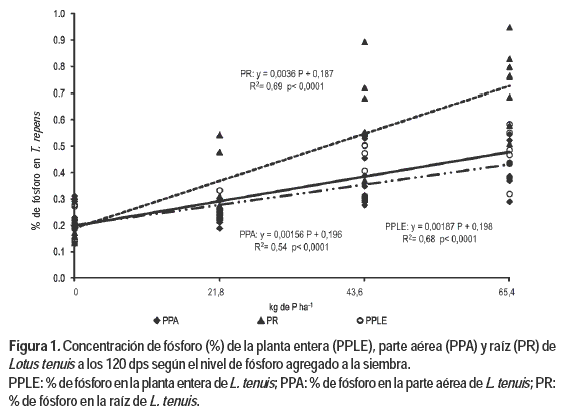
La fertilización fosfatada a la siembra aumentó la concentración de este nutriente en el suelo y la absorción de este por parte de las plántulas de las leguminosas sembradas (Ayala Torales et al., 1998). El aumento del contenido de P al nivel radicular en las leguminosas ocurre por una mayor eficiencia de absorción por unidad de raíz, lo cual compensó el bajo volumen del suelo que exploran y el mayor requerimiento que le imponen los nódulos (Ayala Torales et al., 1998; Puig y Ferrando, 1983; Caradus, 1980).
Los resultados obtenidos por estos autores son coincidentes con los derivados de este estudio en lo referente a la capacidad del L. tenuis de aumentar el contenido de P en el tejido radical por el incremento de este nutriente en el suelo, producido por el incremento en los niveles de P aplicado.
La mayor capacidad de absorción de P del Lotus spp. se debe a una mayor extensión de su sistema radicular. Células más grandes en las raíces son capaces de albergar a su vez vacuolas más grandes (sitio de acumulación de P) lo cual pudo haber favorecido un establecimiento temprano de las plántulas, especialmente en suelos con baja disponibilidad de P (Bermúdez et al., 2000; Puig y Ferrando, 1983).
b) Concentración de N en la planta entera (NPLE), la parte aérea (NPA) y raíz (NR)
La concentración de NPLE en el tejido no presenta diferencias significativas (p<0,05) ante un aumento en las dosis de P agregado. Este contenido de NPLE es el resultado de un comportamiento similar observado en el contenido de NPA y NR, los cuales no presentaron diferencias significativas (p<0,05) ante un aumento en las dosis de P agregado.
Sin embargo, el contenido de este nutriente en la materia seca producida de la parte aérea aumentó por un incremento del peso de las plantas ante el aumento en las dosis de P agregado, resultando ello en un mayor valor absoluto de N en la dieta ofrecida (Silveira, 2005; Silveira et al., 2004a). En el Cuadro 1 se presentan los valores promedio para las variables NPLE, NPA y NR.
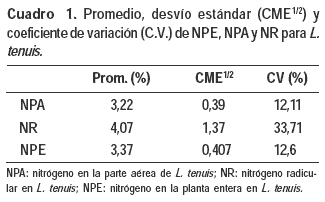
c) Relación N/P en la planta entera (NPLE/PPLE), en la parte aérea (NPA/PPA) y raíz (NR/PR)
La relación NPLE/PPLE presentó una respuesta negativa y significativa frente al incremento de P, ajustando un modelo lineal (Figura 2). Este resultado observado en la relación (NPLE/PPLE) es consecuencia de una respuesta mostrada por sus componentes NPA/PPA y NR/PR, los cuales presentaron una respuesta negativa y significativa frente al aumento en las dosis de P agregado.
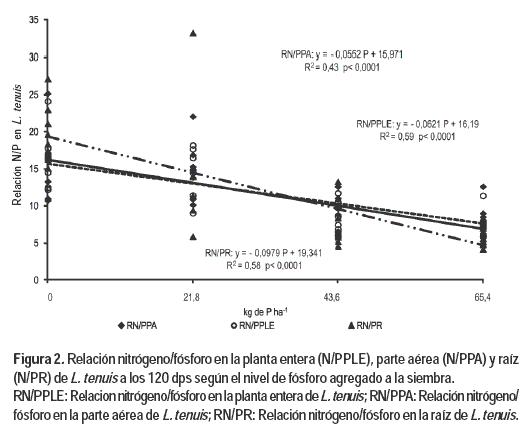
La respuesta negativa de estas relaciones ocurre debido a una acumulación de P en el tejido a medida que se incrementan los niveles de P agregado, mientras que el contenido de N para las distintas fracciones analizadas no presentó diferencias significativas entre los distintos tratamientos (Silveira, 2005; Silveira et al., 2004b). El mayor coeficiente angular de la recta NR/PR se debe a una mayor tasa de acumulación de P realizada por las raíces del L. tenuis las cuales poseen una capacidad de intercambio catiónico del entorno de los 20 meq/100g, además de las características ya mencionadas anteriormente (Marchesi y Elhordoy, 1993).
Trifolium repens L
a) Concentración de P de la planta entera (PPLE), parte aérea (PPA) y raíz (PR)
La concentración de PPLE en T. repens aumentó frente al aumento en las dosis de P agregado a la siembra, ajustando un modelo lineal (Figura 3). El aumento del PPLE es resultado del aumento positivo (p<0,05) que registraron sus componentes PPA y PR los cuales presentaron una respuesta positiva (p<0,05) frente al aumento en las dosis de P.
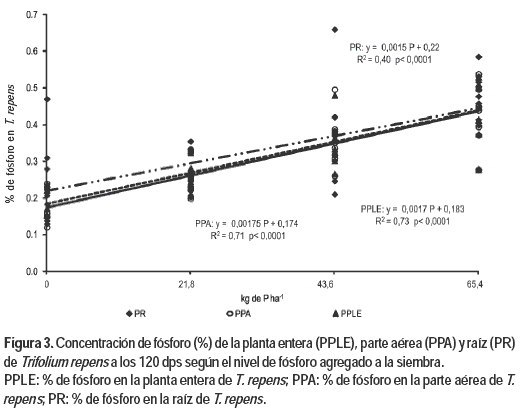
Las concentraciones críticas de nutrientes varían notablemente con las especies, la edad de las plantas, el tejido analizado, las condiciones climáticas registradas durante su crecimiento (Morton et al., 2001; Whitehead, 2000; McNaught y During, 1970), y la etapa de crecimiento de la planta a analizar (Sinclair et al., 1996b). Además, existe un factor estacional que incide en las concentraciones de P en planta y el manejo que se realice de la misma (consumo directo o corte) (Morton y Roberts, 2001; Whitehead, 2000).
Según McNaught y During (1970), la concentración crítica establecida para T. repens en plantas en activo crecimiento y altura de pastoreo es de 0,35 %. Sin embargo, Sinclair et al. (1996b) determinaron diferentes niveles críticos para T. repens en activo crecimiento dependiendo si esta especie se encontraba en tierras bajas (0,43 % P) o en tierras altas y secas (0,22 % P). En cambio Chilibroste et al. (1982) reportaron valores entre 0,32 y 0,36 %, al evaluar la respuesta a cuatro niveles de fertilización a la siembra. En este estudio la concentración de P en planta a los 120 días pos siembra aumentó como respuesta a un mayor nivel de fertilización fosfatada, obteniéndose un valor de 0,46 % para el tratamiento que se aplicó 65,4 kg de P ha-1. La concentración de P en las plántulas correspondiente al nivel máximo de fertilización fue superior al informado por Sinclair et al. (1996b) y Chilibroste et al. (1982) lo cual podría deberse a las diferencias en la etapa fenológica en la cual se realizaron los análisis.
Resultados reportados por Crush (1995), sostienen que la concentración de P en los tejidos de estolones, pecíolos y hojas de T. repens se incrementó como consecuencia de incrementos en la aplicación creciente de fertilizantes fosfatados, siendo estos resultados concordantes con los obtenidos por Olmos (2001) y los registrados en este estudio.
La concentración de P radicular (PR) para T. repens presentó en el modelo propuesto un coeficiente angular muy inferior al observado para L. tenuis lo cual puede ser explicado por el menor desarrollo de la raíz y por no poseer las características morfo-fisiológicas que presenta L. tenuis. Por lo tanto en esta especie se justifica en mayor medida el aporte de la fertilización a la siembra, ya que de lo contrario se estaría afectando en primera instancia el efectivo funcionamiento de la simbiosis rhizobium-leguminosa, la demanda por P de los meristemas, el crecimiento y el rendimiento, y por lo tanto la sobrevivencia de las plantas.
Los resultados obtenidos confirman que para las condiciones particulares de este estudio (tipo de suelo, condiciones ambientales), y en este caso para T. repens cuando un nutriente se encuentra por debajo de los requerimientos para el máximo rendimiento, un aumento en el suministro de P incrementa el crecimiento y el rendimiento, así como la concentración del mismo en la planta.
b) Concentración de nitrógeno de la planta entera (NPE), parte aérea (NPA) y raíz (NR)
En la Figura 4 se presenta la respuesta del NPLE frente al agregado de dosis crecientes de P, donde existen dos zonas que se diferencian en la respuesta al nivel de P agregado. En la primera zona la función de respuesta muestra una tendencia cuadrática con incrementos decrecientes hasta el valor de 38,8 kg de P ha-1, punto a partir del cual no se observó respuesta frente a incrementos en las dosis de P, permaneciendo constante el contenido de NPLE con un valor de 3,18 %.
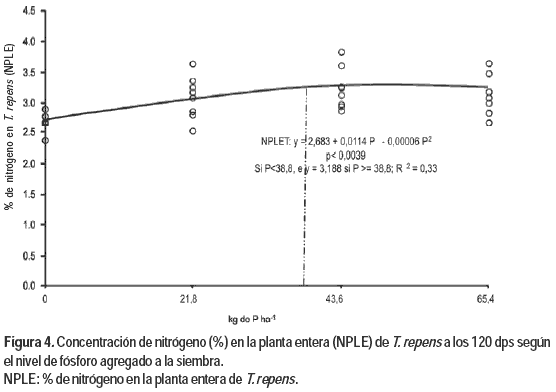
Existe una tendencia a incrementarse el contenido de N en planta frente al agregado de niveles crecientes de P, según lo informan Sinclair et al. (1996b) y McNaught y During (1970). No obstante ello, los valores obtenidos en este estudio resultaron inferiores a los niveles críticos (nivel de suficiencia) determinados en Nueva Zelanda para la producción de forraje en T. repens, a causa de las diferencias en el estado fenológico en el cual se extrajeron las muestras para realizar los análisis (Cornforth y Sinclair, 1982). El valor máximo de N en la parte aérea obtenido en este estudio se ubicó 1,6 % por debajo del nivel crítico de 4,8 % para hoja más pecíolo en estado de activo crecimiento y altura de pastoreo establecido para Nueva Zelanda.
La Figura 5 ilustra la concentración de N en la parte aérea de T. repens (NPA) frente al aumento en las dosis de P. Existen dos zonas distintas en la respuesta al nivel de P agregado. En la primera zona la función presenta una tendencia cuadrática con incrementos decrecientes hasta el valor de 35,6 kg de P ha-1, punto a partir del cual se observa una respuesta constante ante incrementos en las dosis de P, siendo el valor de NPA de 3,18 %.
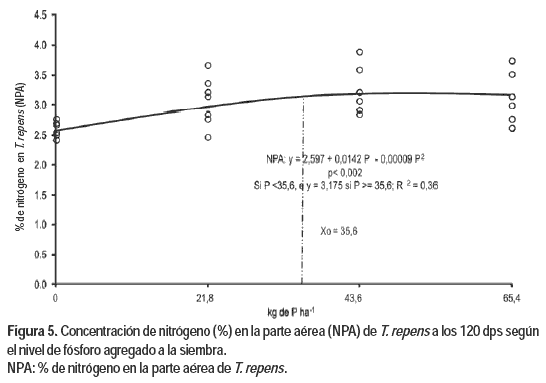
Morón (1999) reporta valores inferiores a este nivel crítico. Las deficiencias en el contenido de N en planta presentaron una significativa asociación con los niveles de P y azufre en planta (Morón, 1999; Sinclair et al., 1996a, 1996b).
La concentración de N en planta se vincula estrechamente con el rendimiento de la pastura y con el incremento en el contenido de proteína en la misma (Olmos, 2001; Morón, 1999).
Resultados obtenidos por Longhurst et al. (1999) en pasturas mixtas (leguminosas y gramíneas) son coincidentes con los registrados en este estudio, ya que las concentraciones de N en la pastura se ubicaron por debajo del óptimo (4,5 %) compatible con el crecimiento de la plantas (Cornforth y Sinclair, 1982). Concluyeron que de acuerdo a los resultados del análisis de planta obtenidos, el N y el P fueron los nutrientes que más limitaron la producción de forraje. Indicaron a su vez, que el P es el nutriente que más limita un adecuado establecimiento de la pastura, fundamentalmente en suelos con bajo contenido de este nutriente y/o con elevada capacidad de fijación.
El contenido de NR no presentó diferencias significativas (p<0,05) y se mantuvo sin cambios ante un aumento en las dosis de P agregado. En el Cuadro 2 se presentan los valores obtenidos para esta variable. El valor promedio obtenido en el presente estudio (3,21 %) se asemeja a los valores reportados por Whitehead (2000) para T. repens en activo crecimiento (2 y 4 %).

c) Relación nitrógeno/fósforo en la planta entera (NPLE/PPLE), en la parte aérea (NPA/PPA) y raíz (NR/PR).
La relación NPLE/PPLE presentó una respuesta negativa y significativa frente al agregado de P.
La relación NPLE/PPLE es el resultado básicamente de un comportamiento similar al que presentó uno de sus componentes (la relación NPA/PPA), la cual mostró una respuesta negativa y significativa frente al agregado de P. El comportamiento de estas variables puede observarse en la Figura 6.
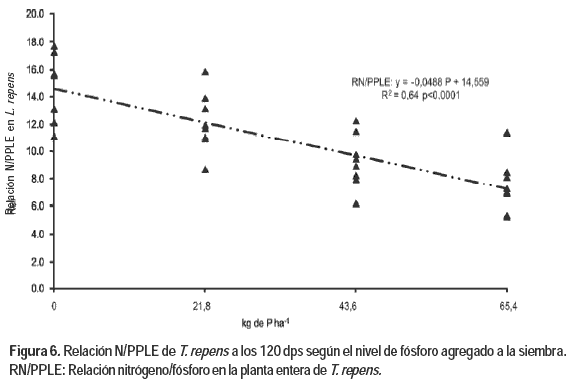
La relación NPA/PPA presentó una tendencia a disminuir con el agregado de niveles crecientes de P, presentándose un valor máximo de 15,28. Resultados obtenidos por McNaught y During (1970) establecen la existencia de deficiencia en P si dicha relación (NPA/PPA) resulta superior a 13 para una pastura en plena producción. Este indicador es solo una guía ya que no se corresponden los resultados a un mismo estado fenológico de las plantas, si resulta evidente que existe una deficiencia de P en T. repens que en este estudio se corresponde con el testigo sin fertilizar. Con una dosis de 21,8 kg de P ha-1 para las condiciones de este estudio esta relación sería inferior a 13 y por lo tanto se esperaría que las plantas guardaran un balance adecuado para producir el 90 % del máximo rendimiento potencial.
La relación NPA/PPA obtenida con el máximo nivel de P agregado (65,4 kg de P ha-1) fue de 7,55, donde se obtuvo la máxima contribución en producción de materia seca de T. repens (Silveira, 2005; Silveira et al., 2004a), este valor obtenido es la mitad de la determinada para Nueva Zelanda en una pastura en crecimiento activo.
La relación NR/PR presentó una respuesta negativa y significativa frente al agregado de P (Figura 7). La relación NR/PR máxima determinada en este estudio coincidió, al igual que en la relación NPA/PPA, con el testigo sin fertilizar, siendo el valor mínimo coincidente con el máximo nivel de P agregado (65,4 kg de P ha-1) obteniéndose una relación de 7,40.
Conclusiones
La concentración de fósforo en las plantas de L. tenuis y T. repens se incrementa en forma lineal ante niveles crecientes de fertilización fosfatada a la siembra, siendo este efecto de mayor importancia en la raíz.
Las especies evaluadas poseen una capacidad diferencial de absorber y utilizar el fósforo del suelo, que se refleja en la concentración en sus tejidos y en las relaciones de estos. La fertilización fosfatada no provocó un cambio en la concentración de nitrógeno a nivel tisular en L. tenuis, en tanto en T. repens presentó una respuesta de tipo cuadrática-plateau pero que presenta un escaso poder predictivo debido a un bajo coeficiente de determinación.
La relación N/P en las plantas de L. tenuis y T. repens frente a aumentos en el nivel de fertilización fosfatada a la siembra disminuye, lo que es explicado por una mayor concentración de fósforo en la materia seca producida.
Agradecimientos
Al Ing. Agr. Yerú Pardiñas y personal de la Estación Experimental Profesor Bernardo Rosengurtt, al personal del Laboratorio de Fertilidad de Suelos de la Facultad de Agronomía por el apoyo recibido y especialmente al Ing. Agr. (MSc) Álvaro Califra.
Bibliografía
Ayala Torales AT, Deregibus VA, Moauro PR. 2000. Differential response of forage legumes to phosphorus application. New Zealand Journal of Agricultural Research, 43: 473 – 480.
Ayala Torales AT, Deregibus VA, Moauro PR. 1998. Phosphorus absorption capacity of Lotus corniculatus and Festuca arundinacea during sward establishment. New Zealand Journal of Agricultural Research, 41: 307 – 312.
Barbazán M, Ferrando M, Zamalvide J. 2007. Estado nutricional de Lotus corniculatus L. en Uruguay. Agrociencia (Uruguay), 11(1): 22 – 34.
Bermúdez R, Carámbula M, Ayala W. 2000. Estudio comparativo de diferentes fuentes y dosis de fósforo sobre el comportamiento productivo de un mejoramiento extensivo con trébol blanco y lotus. En: Jornada anual de producción animal : Resultados experimentales. Montevideo : INIA. (Actividades de Difusión : 225). pp. 17 – 24.
Bordoli JM. 1998. Dinámica de nutrientes y fertilización en siembra directa. En: Curso de Acualización para Egresados de Manejo de la Fertilidad de Suelos en Sistemas Extensivos (Cultivos y Pasturas). Cerro Largo : E.E. de Bañado de Medina. pp. 41 – 46.
Bray RH, Kurtz LT. 1945. Determination of total, organic and available forms of phosphorus in soil. Soil Science, 59: 39 – 45.
Caradus J. 1980. Distinguishing between grass and legume species for efficiency of phosphorus use. New Zealand Journal of Agricultural Research, 23: 75 – 81.
Carámbula M. 2002. Pasturas y forrajes : insumos, implantación y manejo de pasturas. Vol. 2. Montevideo: Hemisferio Sur. 371p.
Carámbula M. 1996. Pasturas Naturales Mejoradas. 1ª ed. Montevideo : Editorial Hemisferio Sur. 524p.
Carámbula M. 1977. Producción y manejo de pasturas sembradas. Montevideo : Hemisferio Sur. 464 p.
Chilibroste JI, Mallarino JL, Pisón P. 1982. Evaluación de los requerimientos de fósforo en la instalación de leguminosas forrajeras [Tesis grado] Montevideo : Facultad de Agronomía. 83p.
Cornforth IS, Sinclair AG. 1982. Fertiliser and lime recommendations for pastures and crops in New Zealand. Wellington : Ministry of Agriculture and Fisheries.
Crush JR. 1995. Phosphorus response of white clover (Trifolium repens L.) genotypes selected for tolerance of aluminium toxicity. New Zealand Journal of Agricultural Research, 38: 451 – 456.
DSF. 1976. Carta de Suelos del Uruguay Escala 1:1.000.000. Montevideo : MGAP.
Frame J, Charlton JFL, Laidlaw AS. 1998. Temperate Forage Legumes. Wallingford : CAB International. pp. 245- 271.
Hart AL. 1989. Nodule phosphorus and nodule activity in white clover. New Zealand Journal of Agricultural Research, 32: 145- 149.
Hernández J. 1999. Fósforo. Montevideo : Facultad de Agronomía. 89p.
Kottek M, Grieser J, Beck C, Rudolf B, Rubel F. 2006. World Map of the Köppen-Geiger climate classification updated. Meteorologische Zeitschrift, 15(3): 259 – 263.
Longhurst RD, O’Connor MB, Toxopeus MRJ. 1999. Pasture establishment and fertilizer requirements on rehabilitated land after opencast coal mining in New Zealand. New Zealand Journal of Agricultural Research, 42: 27 – 36.
Marchesi CE, Elhordoy JA. 1993. Limitantes nutricionales para la producción de pasturas mejoradas, efecto de la dosis, fuente de fósforo y encalado en suelos de las Unidades Arroyo Blanco y Zapallar [Tesis de grado]. Montevideo : Facultad de Agronomía. 260p.
McNaught KJ, During C. 1970. Relation between nutrient concentration in plant tissues and responses of white clover to fertilizers on a gley podozol near Westport. New Zealand Journal of Agricultural Research, 13: 567 – 590.
Millot JC, Methol R, Risso D. 1987. Relevamiento de Pasturas Naturales y Mejoramientos Extensivos en el Áreas Ganaderas del Uruguay. Montevideo : FUCREA, CHPA. 199p.
Morón A. 1999. Relevamiento del estado nutricional y la fertilidad del suelo en cultivos de trébol blanco. En: Jornada de trébol blanco. Montevideo : INIA. (Actividades de Difusión ; 200). pp. 1 – 14.
Morton JD, Smith LC, Dodds KG. 2001. Balanced and adequate potassium and phosphorus nutrition of pasture. New Zealand Journal of Agricultural Research, 44: 269 – 277.
Morton JD, Roberts AHC. 2001. Pasture response to soil phosphorus levels measured under mowing and dairy grazing. New Zealand Journal of Agricultural Research, 44: 259 – 268.
Murphy J, Riley HP. 1962. A modified single solution method for the determination of phosphate in natural waters. Analytica Chimica Acta, 27: 31 – 36.
Olmos F. 2001. Mejoramientos de pasturas con Lotus en la región noreste. Montevideo: INIA. (Serie Técnica ; 124). 48p.
Puig A, Ferrando A. 1983. Requerimientos de fósforo en trébol blanco, lotus y trébol carretilla implantados puros y en mezcla [Tesis de grado]. Montevideo : Facultad de Agronomía. 192p.
Scott WR. 1981. Nutrición de la planta de pradera y ciclaje de nutrientes. En: Langer RHM [Eds.]. Las pasturas y sus plantas. Montevideo : Hemisferio Sur. pp. 185 – 208.
Silveira D. 2005. Efecto de la fertilización fosfatada sobre la implantación, producción inicial y composición química de Lotus glaber Mill y Trifolium repens L. sembradas en cobertura [Tesis de grado]. Montevideo : Facultad de Agronomía. 137p.
Silveira D, Vaz A, Boggiano P, Zanoniani R, Hernández J, del Pino A, Cadenazzi M. 2004a. Efecto del nivel de fertilización fosfatada a la siembra sobre el peso de plantas de Lotus glaber Mill. y Trifolium repens L. En: XX Reunión del Grupo Técnico Regional del Cono sur en Mejoramiento y utilización de los recursos forrajeros del area tropical y subtropical; setiembre 2004; Salto, Uruguay. pp. 289-291.
Silveira D, Vaz A, Boggiano P, Zanoniani R, Hernández J, del Pino A, Cadenazzi M. 2004b. Efecto del nivel de fertilización fosfatada a la siembra sobre la composición química de plantas de Lotus glaber Mill. y Trifolium repens L. En: XX Reunión del Grupo Técnico Regional del Cono sur en Mejoramiento y utilización de los recursos forrajeros del área tropical y subtropical; setiembre 2004; Salto, Uruguay. pp. 292 – 293.
Sinclair AG, Morrison JD, Smith LC, Dodds KG. 1996a. Effects and interactions of phosphorus and sulphur on a mown white clover/ryegrass sward : 1. Herbage dry matter production and balanced nutrition. New Zealand Journal of Agricultural Research, 39: 421 – 433.
Sinclair AG, Morrison JD, Smith LC, Dodds KG. 1996b. Effects and interactions of phosphorus and sulphur on a mown white clover/ryegrass sward : 2. Concentrations and ratios of phosphorus, sulphur, and nitrogen in clover herbage in relation to balanced plant nutrition. New Zealand Journal of Agricultural Research, 39: 435- 445.
Wedderburn ME, Adam KD, Greaves LA, Carter JL. 1996. Effect of oversown ryegrass (Lolium perenne) and white clover (Trifolium repens) on the genetic structure of New Zealand hill pastures. New Zealand Journal of Agricultural Research, 39: 41 – 52.
Whitehead DC. 2000. Nurtient elements in grassland : soil-plant-animal relationships. Wallingford : CAB International. 369p.

