



![Determination of Some Quality Attributes of Feijoa Fruits [Acca sellowiana (Berg) Burret] at Different Ripening Stages](/img/en/prev.gif)






Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
Related links
Share

 Permalink
PermalinkAgrociencia (Uruguay)
Print version ISSN 1510-0839On-line version ISSN 2301-1548
Agrociencia Uruguay vol.19 no.1 Montevideo June 2015
Cobertura del suelo con cal: efecto en la reducción de la temperatura y cambio de color de mandarinas precoces
Gambetta Giuliana1, Mesejo Carlos2, Gravina Alfredo1, Agustí Manuel2, Fasiolo Carolina1, Rey Florencia1, Reig Carmina2, Martínez-Fuentes Amparo2, Bentancur Oscar1
1Facultad de Agronomía, Universidad de la República. Garzón 780, 12900 Montevideo, Uruguay. Correo electrónico: gambetta@fagro.edu.uy
2Instituto Agroforestal Mediterráneo, Universidad Politécnica de Valencia. Camino de Vera s/n, 46022, Valencia, España.
Recibido: 25/6/14 Aceptado: 12/12/14
Resumen
El color de los frutos cítricos es una característica importante para su comercialización. En la medida en que la maduración interna y externa presentan una regulación relativamente independiente, en las mandarinas precoces la cosecha de los frutos se realiza sin haber alcanzado la coloración adecuada para su comercialización. Considerando que el cambio de color de los frutos cítricos se asocia a la disminución de la temperatura, en este trabajo se evaluó la eficacia de la cobertura del suelo con cal en el adelanto del cambio de color de mandarinas Satsuma y Clementina. Los experimentos se realizaron en España (Satsuma 'Iwasaki' y Clementina 'Clemenpons', injertados sobre citrange 'Carrizo') y en Uruguay (Satsuma 'Okitsu' y 'Clementina de Nules', injertados sobre P. trifoliata). La cal se aplicó entre los dos y cuatro meses previos a la cosecha. El tratamiento disminuyó la temperatura del suelo entre 0,5 °C y 3 °C en todas las situaciones. En Satsuma el tratamiento no logró anticipar el cambio de color, ya que la temperatura del suelo permaneció por encima de 23 °C y 18 °C, umbrales por debajo de los cuales disminuye la actividad de las raíces de citrange 'Carrizo' y P. trifoliata. En Clementina, que madura dos meses más tarde, la temperatura del suelo permaneció por debajo de los umbrales durante por lo menos un mes antes de la cosecha, adelantando significativamente la coloración de los frutos y permitiendo incrementar el porcentaje cosechado en forma temprana.
Palabras clave: CLEMENTINA, SATSUMA, DESVERDIZADO, MADURACIÓN
Summary
Hydrated Lime Soil Coverage: Effect On Soil Temperature Reduction And Early Mandarin Colour Break
Citrus fruit colour is an important marketing feature. In view of the relative independence between internal and external maturity, in early mandarins fruit harvest takes place before reaching fruit full colouration. Since citrus fruit colour break is associated to natural temperature reduction, this work evaluated the efficiency of hydrated lime soil coverage on the advance of fruit colour break of Satsuma and Clementine. Experiments were carried out in Spain (Satsuma 'Iwasaki' and Clementina 'Clemenpons', grafted on citrange 'Carrizo') and in Uruguay (Satsuma 'Okitsu' and 'Clementina de Nules', grafted on P. trifoliata). Hydrated lime was applied between two and four months before harvest. Treatment diminished soil temperature between 0.5 °C and 3 °C in all situations. In Satsuma, treatment could not advance colour break, since soil temperature remained above 23 °C and 18 °C, thresholds below which root metabolism of citrange 'Carrizo' and P. trifoliata, respectively, is reduced. In Clementine, maturing two months later, the soil temperature remained below thresholds during at least one month before harvest, significantly advancing fruit colour break and increasing the percentage of fruit harvested early.
Keywords: CLEMENTINE, SATSUMA, DEGREENING, MATURATION
Introducción
El comercio mundial de frutos cítricos para el consumo en fresco exige la obtención de productos de alta calidad. El concepto de calidad comprende por un lado la madurez interna, que involucra fundamentalmente el porcentaje de jugo, el índice de madurez (relación azúcares: ácidos) y la ausencia de semillas, y por otro lado, las características externas, entre las cuales el tamaño, la finura de la piel y el color son las más apreciadas. En los cítricos, la pulpa y la piel de los frutos se desarrollan de forma relativamente independiente (Monselise, 1977), por lo que las mandarinas de maduración precoz alcanzan los estándares de maduración interna antes de lograr la coloración externa requerida. Esto genera la necesidad de desverdizarlas con etileno, hormona que reprime la biosíntesis de clorofila (Fujii et al., 2007), promueve su degradación (Shimokawa et al., 1978; Trebitsh et al., 1993; Katz et al., 2004), y estimula la síntesis de carotenoides (Rodrigo et al., 2006; Fujii et al., 2007; Rodrigo y Zacarías 2007), pero no afecta la madurez interna (Mayuoni et al., 2011). Sin embargo, su aplicación acelera la senescencia de los frutos y su pérdida de calidad (Eckert y Eaks, 1989; Plaza et al., 2004), e incrementa la incidencia de 'rind breakdown' en 'Clementina de Nules' (Cronje et al., 2011). Adicionalmente, el tratamiento en las cámaras aumenta los costos de producción.
Nutricionalmente, la toma de color de los frutos cítricos se ha asociado al incremento de carbohidratos en el flavedo y, sobre todo, de los azúcares reductores (Huff, 1984; Sala et al., 1992; Fidelibus et al., 2008; Gambetta et al., 2012). El aumento de la concentración de carbohidratos se debe, además, al incremento del transporte de sacarosa (Holland et al., 1999) y probablemente también al aumento del gradiente de sacarosa desde el albedo hacia el flavedo (Fidelibus et al., 2008). Asimismo, la hidrólisis de la sacarosa se incrementa en el flavedo de los frutos durante la maduración (Holland et al., 1999).
El metabolismo del nitrógeno está involucrado negativamente en la toma de color de los frutos cítricos (Huff, 1984), de modo que su concentración en el flavedo disminuye durante el cambio de color (Iglesias et al., 2001; Gambetta et al., 2012), y la aplicación de nitratos lo retrasa (Jones y Embleton, 1959; Huff, 1983; Lee y Chapman, 1988; Sala et al., 1992), inhibiendo la degradación de clorofilas y retrasando la síntesis de carotenoides (Alós et al., 2006).
El cambio de color de los frutos cítricos se asocia al descenso de la temperatura del aire y del suelo (Erickson, 1960; Young y Erickson, 1961; Soonen et al., 1979). El cero biológico de los cítricos se sitúa en el entorno de los 13 °C. La elongación de la raíz de diferentes portainjertos se detiene con temperaturas inferiores a 13 ºC (Castle, 1978) y se correlaciona linealmente con la temperatura del suelo entre 18 ºC y 28 ºC (Bevington y Castle, 1985). En citrange 'Carrizo' su crecimiento se reduce al mínimo por debajo de 22 °C (Bevington y Castle, 1985), mientras que el umbral para la detención del desarrollo y metabolismo de las raíces de P. trifoliata se sitúa en 15 °C (Poerwanto et al., 1989).
Las primeras evidencias que relacionan la disminución de la temperatura de las raíces con la coloración de los frutos cítricos se obtuvieron en condiciones controladas. En frutos de 'Calamondín' (Citrus madurensis Lour.) y mandarino Satsuma, el contenido de carotenoides del flavedo se duplicó con una disminución de la temperatura de 30 ºC a 14 ºC (Soonen et al., 1979), y en naranjo dulce 'Valencia' la menor concentración de clorofilas y la mayor concentración de xantofilas en el flavedo, se logró con una temperatura diurna de 20 °C, nocturna de 7 ºC y con una temperatura del suelo de 12 °C (Young y Erickson, 1961).
La reducción de la temperatura inhibe la translocación de sustancias hormonales desde las raíces (Eilati et al., 1969), disminuye la concentración de GA1 en la parte aérea de las plantas (Vidal et al., 2003) y reduce la absorción (Chapman y Parker, 1942) y translocación de nitrógeno (Wallace, 1953). Asimismo, el descenso de la temperatura favorece el aporte de azúcares reductores a los frutos, promoviendo la actividad de la invertasa tanto en los frutos que permanecen en el árbol (Purvis y Rice, 1983), como en aquellos expuestos a bajas temperaturas luego de la cosecha (Hyde et al., 1999). Como consecuencia de todo lo anterior, el crecimiento vegetativo disminuye, aumentando la disponibilidad de carbohidratos para los frutos, lo que promueve el cambio de color (Huff, 1983; Goldschmidt, 1988).
La reducción de la temperatura del suelo con mallas plásticas blancas, colocadas debajo de los árboles, aumentó el tiempo de exposición de las raíces entre 20 °C y 23 °C durante los dos meses previos a la recolección. Como consecuencia, adelantó el cambio de color de los frutos de mandarino 'Clemenpons' injertado sobre citrange 'Carrizo', incrementando el porcentaje de frutos cosechados en la primera fecha de recolección (Mesejo et al., 2012). Adelantar el cambio de color de los frutos de variedades precoces y/o mejorar la intensidad de su coloración, permitiendo anticipar su recolección, disminuir el tiempo de desverdizado y, consecuentemente, mantener la calidad de los frutos durante la post-cosecha, tendría un impacto positivo en la rentabilidad. La utilización de mallas propuesta por Mesejo et al. (2012) resulta dificultosa, por lo que una alternativa de aplicación agronómica, sería contar con materiales capaces de disminuir la temperatura del suelo en igual o mayor magnitud que las mallas plásticas y que al mismo tiempo fueran más económicos y de fácil manejo. La cobertura del suelo con cal [Ca (OH)2] puede ser una opción, sobre todo en plantaciones de suelo ácido. El objetivo del trabajo fue evaluar la eficacia de la cobertura del suelo con cal, en la reducción de la temperatura y su efecto en el adelanto del cambio de color de los frutos de mandarinos de maduración precoz, en plantaciones comerciales de España y Uruguay.
Materiales y métodos
España
Los experimentos se llevaron a cabo en una plantación comercial ubicada en Huelva (37° 24' N, 07° 03' O), en mandarino Clementina 'Clemenpons' (Citrus clementina Hort. ex Tan.) y en mandarino Satsuma 'Iwasaki' (Citrus unshiu Mark.), injertados sobre citrange 'Carrizo' [Citrus sinensis (L.) Osb. × Poncirus trifoliata (L.) Raf.], durante tres años y un año, respectivamente. La distancia de plantación era 6,5 m x 3 m, con orientación N-S y un diámetro promedio de copa de 3,0 m (Clementina) y 3,5 m (Satsuma). Los experimentos se desarrollaron en un suelo limo-arenoso, de pH 6,3, con camellones y riego localizado. Se aplicaron dos tratamientos: control (suelo desnudo en la fila) y cobertura con cal [Ca (OH2)] cubriendo la superficie del terreno por debajo de los árboles 1,5 m desde el tronco hacia cada lado de la entrefila, quedando 3,5 m de ancho para la circulación de maquinaria. El tratamiento se aplicó al inicio del verano, manteniéndose durante los cuatro meses previos a la cosecha en Clementina (julio a noviembre), y dos meses en Satsuma (julio a setiembre). El encalado se mantuvo durante todo el período, realizando en caso de ser necesario, la reposición del material luego de las precipitaciones; el total de cal aplicada fue en promedio 4,5 kg m-2. Se utilizó un diseño de bloques al azar con cuatro repeticiones de cinco árboles cada una.
Uruguay
Se realizaron tres experimentos, dos en plantaciones de mandarino 'Clementina de Nules' localizadas en San José (34° 64' S, 56° 61' O) y Paysandú (31° 38' S, 57° 48' O) y uno en una plantación de mandarino Satsuma 'Okitsu', situada en Salto (31° 38'S, 57° 83' O). En Clementina, la orientación de las filas era N-S, el marco de plantación 5,5 m x 3 m y 7 m x 4 m y el diámetro promedio de las copas de 1,50 m y 2,70 m, en San José y Paysandú respectivamente. En Satsuma, la orientación de las filas era E-O, la distancia de plantación 4 m x 3 m y el diámetro promedio de la copa 2,40 m. En todos los casos el portainjertos utilizado fue P. trifoliata. Los experimentos se desarrollaron en suelos limo-arcillosos, con pH 5,2-5,5, sin camellones y con riego localizado. Se aplicaron los mismos tratamientos que en España, durante los tres meses previos a la cosecha (febrero a mayo en Clementina y diciembre a marzo en Satsuma). En promedio el total de cal aplicada fue 1,3 kg m-2 en Satsuma y 1,5 kg m-2 en Clementina. Se utilizó un diseño de parcelas al azar con cuatro repeticiones de un árbol cada una.
En ambos países la fertilización, el manejo de plagas y enfermedades y la poda se realizaron de acuerdo a las prácticas habituales de producción.
Evaluaciones
En todos los casos se registró la temperatura del aire en la copa de los árboles y del suelo a 20-30 cm de profundidad, dado que la mayor densidad de raíces se encontraba entre 10 y 40 cm, utilizando sensores Thermochron® (iButton DS-1921G-F5, Dallas Semiconductors, EUA), programados para el registro horario de la temperatura. Se estudió la evolución de la temperatura máxima, media y mínima, se contabilizó la proporción de horas con valores por debajo de umbrales asociados a la disminución del metabolismo de los cítricos (23 °C, 22 °C, 18 °C, 15 °C, 13 °C) y la proporción de días con amplitud térmica mayor a 10 °C.
Desde el comienzo del experimento hasta la cosecha, se midió quincenalmente el color de 10-15 frutos por árbol, seleccionados al azar a una altura de 1,5 m sobre el suelo, con un colorímetro digital (Color Reader CR-300 y CR-10, Minolta Co, Ltd, Japan, en España y Uruguay, respectivamente), efectuando 2-3 mediciones en la zona ecuatorial del fruto, utilizando el sistema Hunter Lab en España y CIE-Lab en Uruguay. Los resultados se presentan en el sistema Hunter Lab; se considera cambio de color, cuando la variable a y por lo tanto la relación a/b pasan de negativo a positivo. La primera cosecha se realizó seleccionando los frutos que habían alcanzado el color estándar mínimo para iniciar el tratamiento de desverdizado (a/b entre -0,01 y -0,10 en Clementina y entre -0,20 y -0,40 en Satsuma), y en la segunda se recolectaron los frutos restantes. En ambas fechas se contabilizó el número de frutos cosechados por parcela y se tomó una muestra de 10 frutos por repetición para determinar la madurez interna; la concentración de sólidos solubles totales (°Brix) se determinó con un refractómetro digital (Atago, Japan) y la acidez titulable (%) por valoración con NaOH (0,1 N) utilizando fenolftaleína (1 %) como indicador.
En Uruguay para evaluar el posible efecto de la cal en el pH del suelo, en cada parcela se tomaron muestras a dos profundidades: 0-5 cm y 5-15 cm, al inicio del experimento y un año después; se secaron en estufa a 40 °C durante 48 horas y se molieron hasta un tamaño menor a 2 mm. El pH en agua se determinó por potenciometría (relación suelo/agua = 1/2,5).
El efecto de los tratamientos de campo sobre el tiempo de desverdizado se determinó en la fruta proveniente de las dos fechas de cosechas de los experimentos de Paysandú y Salto (Uruguay). El desverdizado se realizó en una cámara con concentración de 1-3 mg l-1 de etileno, CO2<0,2 %, a 18-22 °C y 90-95 % de humedad relativa (HR) y se contabilizaron las horas necesarias para alcanzar el estándar de color requerido para la comercialización del cultivar (a/b ³ 0,50 en Clementina y a/b ³ 0,10 en Satsuma). Finalizado el desverdizado, se almacenó en frío una caja de 10 kg por parcela durante 30 días a temperaturas entre 4-6 °C, HR>85 % y CO2<0,5 %, y, posteriormente, se trasladó a temperatura ambiente, simulando el transporte desde Uruguay hasta la Unión Europea, principal destino de la fruta. A los 10 días, en una muestra de 50-60 frutos por caja se evaluó el color, el aspecto del cáliz (proporción de frutos con cáliz de color verde, amarillo, marrón o ausente) y el porcentaje de frutos manchados o afectados por podredumbres (incidencia).
Análisis estadísticos
Las variables continuas se analizaron ajustando modelos lineales generales (ANAVA) y la separación de medias se hizo con prueba Tukey (p ≤ 0,05); las proporciones se analizaron ajustando modelos de distribución binomial con función de enlace logit {Ln [p/(1-p)]} con pruebas de razón de verosimilitud.
Resultados
España
En los tres años de experimentos realizados en Clementina 'Clemenpons', la temperatura media del aire se redujo progresivamente de 25,5 °C a 13,5 °C entre el mes de julio y el mes de noviembre; la amplitud térmica diaria también disminuyó de 15,7 °C a 10,6 °C en dicho período. El color de los frutos permaneció prácticamente estable hasta mediados de setiembre, momento a partir del cual el cambio de color evolucionó paralelamente a la disminución de la temperatura. El primer año, a fines de octubre, los frutos control presentaron una relación a/b de -0,35, alcanzando el cambio de color a mediados de noviembre (Figura 1), luego de dos meses con una temperatura media del aire por debajo de 22 °C durante más del 70 % de los días. La amplitud térmica diaria, sin embargo, fue superior a 10 °C entre el 73 % y el 100 % de los días de todo el período (los 31 días de julio y agosto, 22 días de setiembre y 24 de octubre y noviembre). La temperatura media del suelo disminuyó entre 26 °C y 16 °C desde el inicio del experimento hasta la última cosecha. La temperatura máxima promedio alcanzó 29 °C en el mes de julio y disminuyó a 25 °C durante agosto. En la primera quincena de setiembre, dos meses antes del cambio de color, se redujo a 22 °C y el último mes (desde mediados de octubre), disminuyó a 18 °C. Los dos años siguientes se registraron valores similares (datos no presentados).
La cobertura con cal logró disminuir la temperatura media del suelo entre 0,5 °C y 1 °C respecto al control, fundamentalmente debido a la reducción de la temperatura máxima diaria del suelo. Las mayores diferencias entre tratamientos se establecieron a partir de setiembre; en este mes, el primer año de experimentos, la cobertura con cal incrementó un 20 % el tiempo de permanencia por debajo de 23 °C. Paralelamente, el tratamiento de cobertura del suelo modificó la evolución del color de los frutos, presentando significativamente mayor relación a/b desde fines de setiembre y logrando el cambio de color quince días antes (30 de octubre) que los frutos control (15 de noviembre) (Figura 1). A mediados de noviembre, en el momento de la cosecha final, los frutos restantes, provenientes de los árboles con cobertura en el suelo, presentaron una coloración más intensa que los frutos control (a/b =+0,28 y 0, respectivamente). Resultados similares se obtuvieron en los otros dos años de experimentos, aunque con ligeras diferencias en las fechas (datos no presentados).
En los tres años, el adelanto de la coloración de los frutos con el tratamiento de cobertura del suelo permitió aumentar significativamente el porcentaje de frutos cosechados en la primera fecha de recolección (Cuadro 1). El peso medio del fruto y la madurez interna no fueron afectados por el tratamiento (datos no presentados).

En el experimento de Satsuma, de maduración más temprana, la temperatura media del suelo durante el período evaluado fue de 24,8 °C. La cobertura con cal, al igual que en el experimento de Clementina, la redujo entre 0,5 °C y 1 °C. Sin embargo, al tratarse de un período más cálido (15 de julio-13 de setiembre), solo permaneció por debajo del umbral (23 °C) un 7 % del tiempo en el suelo control y un 12 % en el suelo cubierto con cal. Como consecuencia, no se logró adelantar significativamente el cambio de color de los frutos ni el porcentaje de frutos recolectados en la primera fecha (13 de setiembre), cosechándose el 12 % de los frutos provenientes de árboles control y el 14 % del tratamiento de cal. El peso medio de los frutos y la madurez interna tampoco fueron afectados por el tratamiento (datos no presentados).
Uruguay
En los experimentos realizados en 'Clementina de Nules', la temperatura media del aire se redujo desde el inicio del experimento hasta la cosecha, de 21,5 °C a 11,3 °C en San José (marzo a mayo), y de 24,3 °C a 15,0 °C en Paysandú (febrero a mayo). A partir de abril permaneció por debajo de 18 °C durante más del 70 % del tiempo en ambas zonas. La amplitud térmica diaria se mantuvo estable durante todo el período, siendo en promedio de 13 °C en San José y de 15,5 °C en Paysandú, superando los 10 °C durante más del 60 % de los días.
En San José, la temperatura media del suelo varió entre 21,7 °C y 13,4 °C desde el inicio del experimento hasta la cosecha y en Paysandú entre 23,1 °C y 15,6 °C, respectivamente. La cobertura con cal logró disminuirla entre 1 °C y 2,5 °C respecto al control. En San José dicho efecto se dio durante todo el período evaluado (Figura 2), mientras que en Paysandú, el tratamiento solo permitió reducir la temperatura del suelo a partir de abril (datos no presentados).
La disminución de la temperatura del suelo con la cobertura, permitió que este estuviera más tiempo por debajo de 18 °C, 15 °C y 13 °C, umbrales relacionados con la disminución del metabolismo de los cítricos y sus portainjertos. Las diferencias importantes entre tratamientos se observaron a partir de abril (Cuadro 2), momento clave en la determinación del cambio de color. En ambas zonas, la cobertura del suelo permitió acumular un mayor porcentaje de horas con temperatura por debajo de 18 °C. Sin embargo, en ambos experimentos fue muy escasa la acumulación de horas por debajo de 15 °C, temperatura que provoca el cese del desarrollo y actividad radical del portainjerto P. trifoliata, o por debajo de 13 °C, cero biológico de los cítricos. La mayor acumulación por debajo de 13 °C se verificó en el mes de mayo, cuando ya más del 50 % de los frutos habían cambiado de color (Cuadro 2).
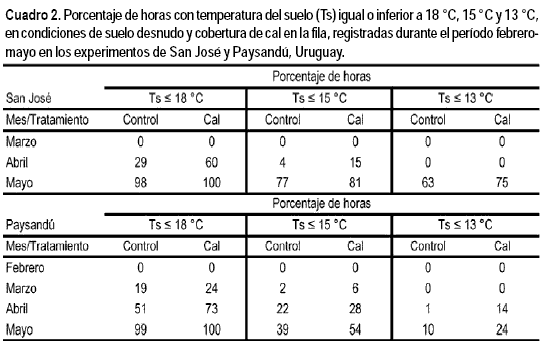
El color de los frutos prácticamente no varió hasta la primera quincena de abril. A partir de ese momento, la relación a/b se incrementó notablemente en respuesta a la disminución de la temperatura. En la Figura 3 se presenta la evolución del color de los frutos del experimento de Paysandú. En la primera fecha de recolección (2 mayo), los frutos provenientes del tratamiento con cal presentaban una relación a/b mayor que los controles (-0,30 y -0,41, respectivamente). Valores similares, aunque sin alcanzar diferencias significativas, se obtuvieron en el experimento de San José (datos no presentados).
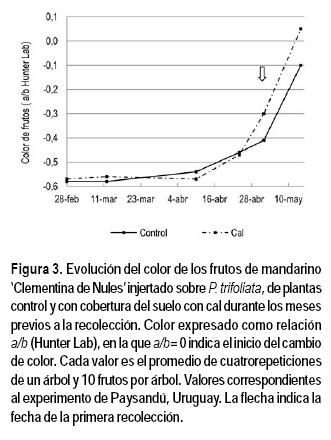
En ambos experimentos la cobertura del suelo con cal incrementó significativamente el porcentaje de frutos recolectados en la primera fecha (Cuadro 3). Tampoco en los experimentos de Uruguay se modificó el peso medio del fruto ni la madurez interna (datos no presentados). Al igual que en España, en la cosecha final, en la que se recolectaron los frutos restantes, los provenientes del tratamiento de cal presentaron coloración significativamente más intensa que los de las plantas control, alcanzando una relación a/b de 0,05 y -0,10, respectivamente (Figura 3). En San José, los valores fueron similares, aunque sin alcanzar diferencias significativas (datos no presentados).

En el experimento de Satsuma 'Okitsu', realizado en Salto entre diciembre y marzo, la temperatura del aire y del suelo fue más elevada que en los experimentos de 'Clementina de Nules'; en dicho período, la temperatura del aire disminuyó, en promedio, de 30 °C a 22 °C. El tratamiento de cobertura con cal permitió disminuir aproximadamente 3 °C la temperatura promedio del suelo en relación al control. Durante las dos semanas previas a la cosecha, se incrementó el porcentaje de horas de permanencia del suelo por debajo de 22 °C (67% y 27 %, respectivamente), aunque no disminuyó por debajo de 18 ºC. En los quince días previos a la primera fecha de recolección el suelo estuvo por debajo de dicho umbral solo un 2 % del tiempo en el control y un 3 % en el cubierto con cal, por lo que no se logró adelantar el cambio de color, ni se modificó la calidad interna de los frutos, al igual que en el experimento de Satsuma realizado en España. En la primera fecha de cosecha (14 de marzo) se recolectó el 13 % de los frutos control y el 15 % de los frutos provenientes del tratamiento con cobertura, sin alcanzar diferencias significativas entre tratamientos. En la cosecha final (28 de marzo), sin embargo, los frutos provenientes del tratamiento de cal presentaron un valor significativamente mayor de la variable b que los controles (b = 25 y 23, respectivamente), indicando una coloración con mayor tonalidad amarilla.
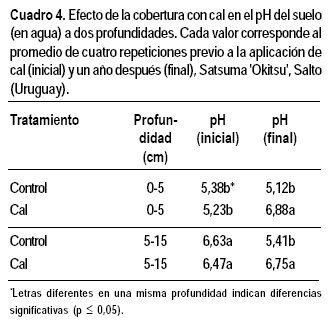
Efecto del tratamiento sobre el pH del suelo
En los tres experimentos, el tratamiento de cal tendió a aumentar el pH del suelo en los primeros 5 cm del perfil, siendo el aumento significativo solo en dos de ellos. A mayor profundidad, la cobertura de cal no modificó el pH inicial (Cuadro 4).
Efecto del tratamiento en el tiempo de desverdizado y calidad del fruto.
En la medida en que la primera cosecha se realizó seleccionando los frutos por color (a/b entre -0,01 y -0,10 en Clementina y -0,20 y -0,40 en Satsuma), el tiempo de desverdizado no varió entre los tratamientos en ninguno de los cultivares. Sin embargo, en la cosecha final, en la que se recolectaban todos los frutos del árbol, los provenientes del tratamiento de cal, que presentaban mejor coloración que los frutos control, requirieron menor tiempo de desverdizado (Cuadro 5). Las horas necesarias variaron notablemente según el cultivar; los frutos de 'Clementina de Nules' necesitaron más de 120 horas de desverdizado para lograr el color requerido para su comercialización (a/b ³ 0,50), mientras que los de Satsuma 'Okitsu' lograron la coloración estándar (a/b ³ 0,10) a partir de las 48 horas de tratamiento.

No se detectaron efectos consistentes del tratamiento de cal en la calidad de los frutos, luego de 30 días de almacenamiento en frío y 10 días de vida de mostrador a temperatura ambiente. El porcentaje de frutos con podredumbres o manchas en la piel fue muy bajo en general, no superando en ninguno de los casos el 2,5 % de los frutos. En Satsuma 'Okitsu', como consecuencia de la reducción del tiempo de desverdizado, el tratamiento de cal incrementó la proporción de frutos con cáliz verde con respecto al control (70 % y 51 %, respectivamente). En 'Clementina de Nules', solo se encontraron diferencias significativas entre las fechas de cosecha, y no hubo interacción fecha x tratamientos; así, los frutos cosechados en primera instancia, que necesitaron en promedio 30 horas menos de desverdizado que los recolectados en segunda fecha, presentaron mejor aspecto de cáliz que estos últimos (41 % y 1 % de los frutos con cáliz verde, respectivamente).
Discusión
El cambio de color de los frutos de dos cultivares de Clementina, evaluado en tres condiciones ambientales (suroeste de España, sur y norte de Uruguay), con portainjertos adaptados a las condiciones de cada país, se asoció a la disminución de la temperatura del aire y del suelo, y fue promovido al reducir artificialmente esta última. En nuestros experimentos la amplitud térmica per se no estimuló la toma de color, como ha sido demostrado para diferentes especies y cultivares (Agustí, 1999).
En estudios previos, la reducción de la temperatura del suelo con mallas plásticas reflectivas colocadas sobre estructuras por debajo de los árboles permitió, en las condiciones productivas de España, adelantar el cambio de color de frutos de mandarino Clementino 'Clemenpons' injertado sobre citrange 'Carrizo', incrementando el porcentaje de frutos cosechados en la primera fecha de recolección (Mesejo et al., 2012). El adelanto se atribuyó al mayor tiempo de exposición de las raíces entre 20 °C y 23 °C durante los dos meses previos a la recolección. En nuestros experimentos realizados en dos cultivares de Clementina, con un material reflectante de más fácil aplicación (cal), la cobertura del suelo también permitió disminuir su temperatura durante los dos-tres meses anteriores al cambio de color, independientemente de la zona de producción (España o Uruguay). El tratamiento de cal aumentó el tiempo de exposición de las raíces a temperaturas inferiores a 23 °C y 18 °C en España y Uruguay, respectivamente, y, consecuentemente, adelantó el cambio de color de los frutos.
En los cultivares de Satsuma 'Iwasaki' y 'Okitsu', sin embargo, el tratamiento no logró anticipar el cambio de color, independientemente del área de cultivo. Estos maduran a finales del verano, y no fue posible durante dicha estación reducir la temperatura del suelo por debajo del rango necesario para disminuir la actividad de las raíces. Similar dificultad en la reducción de la temperatura por debajo del umbral durante el verano, fue reportada por Mesejo et al. (2012) utilizando mallas reflectantes en Clementino 'Clemenpons'.
El efecto de la disminución de la temperatura del suelo en la reducción del metabolismo de las raíces de portainjertos utilizados en cítricos, se ha estudiado en plantas jóvenes y generalmente en condiciones controladas. En este sentido, se establece que el crecimiento de las raíces comienza a darse por encima de 13 °C (Castle, 1978), y se correlaciona positiva y linealmente con temperaturas entre 18 ºC y 28 ºC, pero es muy reducido por debajo de 22 ºC (Bevington y Castle, 1985). Las raíces de naranjo amargo reducen exponencialmente la respiración con la disminución de la temperatura del suelo entre 30 °C y 15 °C (Bryla et al., 2001) y el crecimiento radical de plantas jóvenes de citrange 'Carrizo' se ve limitado con temperaturas de suelo inferiores a 22 °C (Bevington y Castle, 1985). En los experimentos realizados en España utilizando citrange 'Carrizo' como portainjerto, la cobertura con cal, que permitió incrementar el tiempo de exposición de las raíces por debajo de 23 °C, promovió el adelanto del cambio de color de los frutos. En Uruguay, donde se utilizó como portainjerto el P. trifoliata, fue necesario que la temperatura se redujera por debajo de 18 °C para lograr un efecto similar, lo que sugiere que este portainjerto caducifolio necesita temperaturas más bajas para reducir su metabolismo. Estudios realizados en condiciones controladas indican que 15 °C sería el umbral que logra el cese del desarrollo y de la actividad radical del P. trifoliata (Poerwanto et al., 1989).
El inicio de la coloración en respuesta a las bajas temperaturas se atribuye fundamentalmente a la inhibición de la síntesis y translocación de sustancias tipo giberelinas hacia la parte aérea (Eilati et al., 1969), así como a la reducción de la absorción y transporte de nitrógeno y a una mayor disponibilidad de carbohidratos solubles (Agustí, 1999).
En conclusión, nuestros resultados confirman que la cobertura de la superficie del suelo con cal reduce entre 0,5 °C y 3 °C la temperatura del mismo durante los meses previos a la maduración. Cuando esta disminución incrementa el tiempo de exposición de las raíces por debajo de umbrales asociados al cese del metabolismo, se adelanta el cambio de color de los frutos. En las condiciones del suroeste de España y del sur y norte de Uruguay fue posible adelantar la coloración de frutos de Clementina, que maduran a comienzos del otoño, pero no fue posible hacerlo con los de Satsuma, que maduran a fines del verano.
Agradecimientos
A las empresas Agrimarba S.A. (España), Frutícola Libertad S.A., El Repecho, S.A. y Antonio De Souza e hijos (Uruguay). Trabajo parcialmente financiado por el Programa ALßan, becas de alto nivel de la Unión Europea para América Latina (NI E03D15012UR) y la Comisión Sectorial de Investigación Científica (Universidad de la República, Uruguay).
Bibliografía
Agustí M. 1999. Preharvest factors affecting postharvest quality of citrus fruit. En: Schirra M [Ed.]. Advances in Postharvest Diseases and Disorders Control of Citrus Fruit. Trivandrum : Research Signpost. pp. 1 – 34.
Alós E, Cercós M, Rodrigo MJ, Zacarías L, Talón M. 2006. Regulation of color break in Citrus fruits : Changes in pigment profiling and gene expression induced by gibberellins and nitrate, two ripening retardants. Journal of Agricultural and Food Chemestry, 54: 4888 – 4895.
Bevington KB, Castle WS. 1985. Annual root growth pattern of young trees in relation to shoot growth, soil temperature and water content. Journal of the American Society for Horticultural Science, 110: 840 - 845.
Bryla DR, Bouma TJ, Hartmond U, Eissenstat DM. 2001. Influence of temperature and soil drying on respiration of individual roots in citrus : integrating greenhouse observations into a predictive model for the field. Plant Cell and Environment, 24: 781 – 790.
Castle WS. 1978. Citrus root system : their structure, function, growth and relationship to tree performance. En: International Citrus Congress; 15 - 23 agosto; 1978; Sidney, Australia. Griffith : International Society of Citriculture. pp. 62 – 69.
Chapman HD, Parker ER. 1942. Weekly absorption of nitrate by young bearing orange trees growing out of doors in solution cultures. Plant Physiology, 17: 336 – 376.
Cronje PJ, Barry G, Huysamer R. 2011. Postharvest rind breakdown of ‘Nules Clementine’ mandarin is influenced by ethylene application, storage temperature and storage duration. Postharvest Biology and Technology, 60: 192 – 201.
Eckert JW, Eaks IL. 1989. Postharvest disorders and diseases of citrus. En: Reuter W, Calavan EC, Carman GE. [Eds.]. The citrus industry. Vol. 5. Berkeley : University of California Press. pp. 179 – 260.
Eilati SK, Goldschmidt EE, Monselise SP. 1969. Hormonal control of colour change in orange peel. Experientia, 25: 209 – 210.
Erickson LC. 1960. Colour development in Valencia oranges. Proceedings of the American Society for Horticultural Science, 75: 257 – 261.
Fidelibus MW, Koch KE, Davies FS. 2008. Gibberellic acid alters sucrose, hexoses, and their gradients in peel tissues during color break delay in ‘Hamlin’ orange. Journal of the American Society for Horticultural Science, 133: 706 - 767.
Fujii H, Shimada T, Sugiyama A, Nishikawa F, Endo T, Nakano M, Ikoma Y, Shimizu T, Omura M. 2007. Profiling ethylene-responsive genes in mature mandarin fruit using a citrus 22K oligoarray. Plant Science, 173: 340 - 348.
Gambetta G, Martínez-Fuentes A, Bentancur O, Mesejo C, Reig C. Gravina A, Agustí M. 2012. Hormonal and nutritional changes in the flavedo regulating rind colour development in sweet orange [Citrus sinensis (L.) Osb.) Journal of Plant Growth Regulation, 31: 273 – 282.
Goldschmidt EE. 1988. Regulatory aspects of chloro-chromoplast interconversions in senescing Citrus fruit peel. Israel Journal of Botany, 37: 123 - 130.
Holland N, Sala JM, Menezes HC, Lafuente MT. 1999. Carbohydrate content and metabolism as related to maturity and chilling sensitivity of cv. Fortune mandarins. Journal of Agricultural and Food Chemestry, 47: 2513 – 2518.
Huff A. 1984. Sugar regulation of plastid interconversions in the epicarp of Citrus fruit. Plant Physiology, 76: 307 – 312.
Huff A. 1983. Nutritional control of regreening and degreening in Citrus peel segments. Plant Physiology, 73: 243 – 249.
Hyde J, Jurch G, Baldwin E, Echeverría E. 1999. Low temperature induction of acid invertase activity in flavedo tissue of late season grapefruit (Citrus paradisi). Scientia Horticulturae, 80: 49 – 56.
Iglesias DJ, Tadeo FR, Legaz F, Primo-Millo E, Talón M. 2001. In vivo sucrose stimulation of colour change in citrus fruit epicarps : Interactions between nutritional and hormonal signals. Physiologia Plantarum, 112: 244 – 250.
Jones WW, Embleton TW. 1959. The visual effect of nitrogen nutrition on fruit quality of ‘Valencia’ oranges. Proceedings of the American Society for Horticultural Science, 73: 234 – 236.
Katz E, Martínez-Lagunes P, Riov J, Weiss D, Goldschmidt EE. 2004. Molecular and physiological evidence suggests the existence of a system II-like pathway of ethylene production in non-climacteric Citrus fruit. Planta, 219: 243 – 252.
Lee LS, Chapman JC. 1988. Yield and fruit quality responses of Ellendale mandarins to different nitrogen and potassium fertilizer rates. Australian Journal of Experimental Agriculture, 28:143 – 148.
Mayuoni L, Tietel Z, Patil BS, Porat R. 2011. Does ethylene degreening affect internal quality of citrus fruit? Postharvest Biology and Technology, 62: 50 – 58.
Mesejo C, Gambetta G, Gravina A, Martínez-Fuentes A, Reig C, Agustí M. 2012. Relationship between soil temperature and fruit colour development of ‘Clemenpons’ Clementine mandarin (Citrus clementina Hort ex. Tan). Journal of the Science of Food and Agriculture, 92: 520 - 525.
Monselise SP. 1977. Citrus fruit development, endogenous system, and external regulation. En: II International Citrus Congress; 1 - 8 mayo; 1977; Orlando, EE. UU. Florida : International Society of Citriculture. pp. 664 - 668; v. 2.
Plaza P, Sanbruno A, Usall J, Lamarca N, Torres R, Pons J, Viñas I. 2004. Integration of curing treatments with degreening to control the main post-harvest diseases of Clementine mandarins. Postharvest Biology and Technology, 34(1): 29 - 37.
Poerwanto R, Inoue H, Kataoka I. 1989. Effects of temperature on the morphology and physiology of the roots of Trifoliate orange budded with Satsuma mandarin. Journal of the Japanese Society for Horticultural Science, 58(2): 267 – 274.
Purvis AC, Rice JD. 1983. Low-temperature induction of invertase activity in grapefruit flavedo tissue. Pytochemestry, 22: 831 – 834.
Rodrigo MJ, Zacarías L. 2007. Effect of postharvest ethylene treatment on carotenoid accumulation and the expression of carotenoid biosynthetic genes in the flavedo of orange (Citrus sinensis L. Osbeck) fruit. Postharvest Biology and Technology, 43(1): 14 – 22.
Rodrigo MJ, Alquezar B, Zacarías L. 2006. Cloning and characterization of two 9-cis-epoxycarotene dioxygenase genes, differentially regulated during fruit maturation and under stress conditions, from orange (Citrus sinensis L. Osbeck). Journal of Experimental Botany, 57: 633 – 643.
Sala JM, Cuñat P, Collado M, Moncholi V. 1992. Effect on nitrogenous fertilization (quantity and nitrogen form) in precocity of colour change of ‘Navelina’ oranges. En: VII International Citrus Congress; 8 - 13 marzo; 1992; Acireale, Italia. Catania : International Society of Citriculture. pp. 598 - 602; v. 2.
Shimokawa K, Shimada S, Yaeo K. 1978. Ethylene-enhanced chlorophyllase activity during degreening of Satsuma mandarin (Citrus unshiu Marc.). Scientia Horticulturae, 8: 129 – 135.
Soonen HD, Lenz F, Gross J. 1979. Influence of root temperature on carotenoid development in the peel of Citrus unshiu (Mark.) and Citrus madurensis (Lour.). Gartenbauwissenschaft, 44: 49 – 52.
Trebitsh T, Goldschmidt EE, Riov J. 1993. Ethylene induces de novo synthesis of chlorophyllase, a chlorophyll degrading enzyme, in Citrus fruit peel. Proceedings of the National Academy of Sciences of the United States of the America, 90: 9441 – 9445.
Vidal AM, Ben-Cheikh W, Talón M, García-Martínez JL. 2003. Regulation of gibberellin 20-oxidase gene expression and gibberellin content in citrus by temperature and citrus exocortis viroid. Planta, 217(3): 442 – 448.
Wallace A. 1953 Nitrogen absorption and translocation by citrus cuttings at different root temperatures. Proceedings of the American Society for Horticultural Science, 61: 89 – 94.
Young LB, Erickson LC. 1961. Influence of temperature on color change in Valencia orange. Proceedings of the American Society for Horticultural Science, 78: 197 - 200.

