










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Curriculum ScienTI
Curriculum ScienTILinks relacionados
Compartir

 Permalink
PermalinkAgrociencia (Uruguay)
versión impresa ISSN 1510-0839versión On-line ISSN 2301-1548
Agrociencia Uruguay vol.18 no.2 Montevideo dic. 2014
Diversidad vegetal del campo natural de Butia odorata (Barb. Rodr.) Noblick en Uruguay
Rivas Mercedes1, Jaurena Martín2, Gutiérrez Lucía3, Barbieri Rosa Lía4
1Universidad de la República, Facultad de Agronomía/Centro Universitario Regional Este, Departamento de Biología Vegetal. Garzón 780, 12900 Montevideo, Uruguay. Correo electrónico: mrivas@fagro.edu.uy
2INIA Tacuarembó. Ruta 5 km 386, Tacuarembó, Uruguay
3Universidad de la República, Facultad de Agronomía, Departamento de Biometría, Estadística y Computación. Garzón 780, 12900 Montevideo, Uruguay
4Embrapa Clima Temperado. BR 392, km 78, Caixa postal 403, Pelotas, RS, CEP 96010-971, Brasil
Recibido: 18/11/13 Aceptado: 10/9/14
Resumen
Este trabajo fue realizado con el objetivo de caracterizar la diversidad vegetal del campo natural del palmar de Castillos (Rocha, Uruguay), identificar diferencias en la composición botánica dentro y fuera del palmar, determinar las especies que discriminan los dos ambientes y reconocer las potencialidades de los recursos genéticos y servicios ambientales. Mediante el uso del método de muestreo punto cuadrado, aplicado de dos formas (ocho transectas largas y 40 transectas cortas) se determinó la presencia de 70 taxones. Los representantes de la familia Poaceae son el 57,1 % de los taxones, mayoritariamente perennes, con una relación C4/C3 de 1,6. Los análisis de agrupamientos y de componentes principales mostraron diferencias consistentes en la composición florística del estrato herbáceo del campo con palmas y sin palmas. Estas diferencias se atribuyen a la adaptación de las especies a condiciones de sombreado y mayor humedad del suelo, entre las principales: Stenotaphrum secundatum, Pratia hederacea, Juncus spp., Hydrocotyle bonariensis, Paspalum urvillei, Paspalum proliferum y Plantago sp. El palmar provee de bienes de uso directo como forraje, sombra y abrigo para el ganado, producción de miel y recreación activa. Se reporta la utilización de un grupo destacado de recursos fitogenéticos de uso forrajero, medicinal y ornamental.
Palabras clave: BUTIÁ, CAMPOS, RECURSOS FITOGENÉTICOS, SERVICIOS ECOSISTÉMICOS
Summary
Plant Diversity of Natural Grasslands of Butia odorata (Barb. Rodr.) Noblick Palm Groves in Uruguay
This research was conducted to characterize plant diversity of the natural campo in the palm grove of Castillos (Rocha, Uruguay), to identify botanical composition differences inside and outside the palm grove, to determinate the species that discriminate the two environments, and to recognize the potential of genetic resources and environmental services. Using the square point sampling method in two different strategies (eight long transects and 40 short transects) the presence of 70 taxons was determined. The Poaceae family represented the 57,1 % of the taxons, mainly perennial, with a C4/C3 ratio of 1.6. The cluster and principal component analysis showed consistent differences between the floral composition in the herbaceous stratum of the campo with and without palm trees. These differences can be attributed to species adaptation to factors like shading and higher soil moisture; among the main differential: Stenotaphrum secundatum, Pratia hederacea, Juncus spp., Hydrocotyle bonariensis, Paspalum urvillei, Paspalum proliferum y Plantago sp. The palm grove offers direct use goods like forage, shade and shelter for cattle, honey production and active recreation. We also identify an important group of plant genetic resources for forage, medicinal and ornamental use.
Keywords: BUTIÁ, CAMPOS, PLANT GENETIC RESOURCES, ECOSYSTEM SERVICES
Introducción
Los llamados Pastizales del Río de la Plata constituyen una de las principales áreas de pastizales naturales templado-subtropicales del mundo. Comprenden unos 760.000 km2 de campos y pampas del centro de Argentina, sur de Brasil y Uruguay (Soriano, 1991) (Figura 1). Desde el punto de vista biogeográfico se encuentran en la región Neotropical e integran la Provincia Pampeana (Cabrera y Willink, 1973). En este territorio se distinguen siete subdivisiones (Soriano, 1991), cinco de ellas correspondientes a áreas de pampas y dos a áreas de campos. Los denominados campos del norte corresponden al sur de Brasil y norte de Uruguay, mientras que los campos del sur serían exclusivos de Uruguay (Bilenca y Mirraño, 2004). En Brasil, los campos integran el bioma «Pampa», ocupando el 2,1% del territorio nacional y el 63 % del estado de Rio Grande do Sul (IBGE, 2012). La distinción fitogeográfica de los campos ya había sido indicada por Cabrera y Willink (1973) al identificar un distrito Uruguayense dentro de la Provincia Pampeana y por Chebataroff (1951) que distingue seis regiones naturales en Uruguay.
Uruguay se ubica entre los paralelos 30º y 35º de latitud sur, lo que determina una zona de transición climática subtropical-templado, con veranos calientes, inviernos fríos y sin estación seca. La diversidad geológica, geomorfológica y edafológica presentes en el territorio, a la que se suma la confluencia de distintas floras, han originado diversidad de tipos de campos, tanto por su fisonomía como por su composición específica (Rosengurtt, 1943; Millot et al., 1987; Lezama et al., 2011), además de una alta diversidad de especies y recursos fitogenéticos de valor forrajero, medicinal y ornamental (Millot et al., 1987; Berretta et al., 2007; Rivas, 2010). La heterogeneidad de los campos de Uruguay se ha estudiado para la región del basalto (Lezama et al., 2006), las sierras del este, la región centro-sur y la cuenca sedimentaria del noreste (Lezama et al., 2011). Para los campos del sur de Brasil, los antecedentes también señalan la presencia de diversas comunidades campestres (Boldrini, 2006; Overbeck et al., 2007).
En la región este de Uruguay se distinguen diferentes geomorfologías: sierras, colinas, lomadas y llanuras correspondientes a la cuenca de la laguna Merín, la laguna Negra y las lagunas costeras de la cuenca Atlántica (Durán, 1999; PROBIDES, 1999).
Los palmares de Butia odorata (Barb. Rodr.) Noblick (nombre validado recientemente para Butia capitata (Mart.) Becc., por Noblick, 2011) se distribuyen en el sur de Brasil y sureste de Uruguay, generalmente sobre terrenos planos situados por encima del nivel de inundación (llanuras medias). Son formaciones vegetales constituidas por un estrato arbóreo de palmas butiá con densidades que en Uruguay van desde menos de 50 a más de 500 palmas por hectárea, sobre un estrato herbáceo de campo natural (Rivas, 2005; Zaffaroni et al., 2005). Ocupan unas 70.000 hectáreas distribuidas en dos áreas, Castillos y San Luis (Figura 1), formando parte de la Reserva de Biosfera Bañados del Este (PROBIDES, 1999). La tala o el daño a las palmas adultas está prohibida por Ley Nacional desde 1939, sin embargo el manejo productivo que se realiza en los campos de palmar impide la regeneración de la comunidad. En el área de Castillos, el sobrepastoreo continuo del ganado provoca el consumo de los renuevos y su destrucción por pisoteo; mientras que el cultivo de arroz irrigado que se desarrolla en el palmar de San Luis impide la germinación de las semillas y el desarrollo de plántulas. Es así que los individuos que constituyen actualmente los palmares son centenarios y coetáneos, incrementándose las tasas de mortalidad a medida que pasa el tiempo con el consiguiente riesgo de extinción de la comunidad (Rivas, 2005).
Los campos del palmar de butiá, al igual que otros en el país y la región, se encuentran la mayor parte del tiempo sometidos a sobrepastoreo y a fuertes procesos de cambio en el uso de la tierra. La sustitución de los campos naturales por forestación, cultivos agrícolas y minería, se ha intensificado en los últimos años (Laterra y Rivas, 2005; Paruelo et al., 2006; Overbeck et al., 2007; GEO Uruguay, 2008; Rivas, 2010), con el consiguiente incremento del riesgo de pérdida del ecosistema Palmar de butiá y los recursos fitogenéticos del campo natural. Para la conservación in situ y el manejo sostenible de estos campos es necesario conocer y valorizar la biodiversidad tanto del estrato arbóreo como del herbáceo. Este trabajo se enmarca en la necesidad de priorizar la conservación de la biodiversidad de los pastizales (Faber-Langendoen y Josse, 2010), el bioma olvidado en el diseño de las políticas de conservación (Overbeck et al., 2007; Heidenreich, 2009; Bond y Parr, 2010).
Las unidades de vegetación y la composición florística de los campos del palmar de butiá no han sido estudiadas previamente. El objetivo general en el marco del cual se realiza este trabajo es valorizar el ecosistema del palmar de butiá y generar propuestas para su conservación y utilización sostenible. En este contexto, los objetivos de este trabajo son caracterizar la diversidad vegetal específica del campo natural del palmar de butiá de Castillos (Rocha, Uruguay), establecer si existen diferencias en la composición botánica de los campos dentro y fuera del palmar, determinar cuáles son las especies que discriminan y reconocer potencialidades de los recursos fitogenéticos y servicios ecosistémicos.
Materiales y métodos
Área de estudio
El experimento se instaló en el predio de un productor ganadero (S 34º10’06’’ W 53º 55’43’’) en el área del palmar de Butia odorata en Castillos (Rocha, Uruguay) (Figura 1). La densidad promedio de palmas en el sitio varía entre 50 y 250 palmas/ha. El clima de la región es templado a subtropical, con una precipitación media anual de 1150 mm (CV 22 %) y una temperatura media de 6 ºC (promedio en invierno 10,8 ºC y promedio en verano 21,5 ºC) (Instituto Uruguayo de Meteorología, 1996). Los suelos predominantes son argiacuoles típicos y albacualfes típicos. El uso de la tierra es predominante ganadero (bovinos y ovinos) con una carga promedio de 0,9 UG/ha.
Muestreo de vegetación
Con el objetivo de comparar la composición botánica de la pradera natural del palmar y la que se encuentra por fuera del mismo, se delimitaron dos zonas o estratos: con palmar (CP) y sin palmar (SP) en un área aproximada de diez hectáreas (Figura 2). En cada uno de los estratos se relevó la composición botánica de la pradera natural en la primavera de 1999 mediante el uso del método de muestreo Point Quadrat (Daget y Poissonet, 1971) aplicado de dos formas diferentes. En el primer caso se utilizaron cuatro transectas/estrato de 50 metros de longitud. En cada una de ellas se registraron todos los taxones en puntos de contacto cada 50 centímetros, totalizando 100 puntos por transecta. En el segundo caso se utilizaron transectas colocadas en la diagonal de áreas fijas de 5 x 5 m, colocadas sistemáticamente de forma de cubrir los diferentes microambientes. Se trabajó con 20 áreas en cada estrato CP y SP. El relevamiento de las especies se realizó cada 20 centímetros, totalizando 35 puntos por transecta. Cuando la identidad de las especies no pudo ser determinada, la identificación se realizó a nivel de género. Para ambos métodos se calcularon las frecuencias de los taxones y se reunieron en las matrices respectivas. Para cada taxón se revisó la información relativa a ciclo biológico, distribución y adaptación (Rosengurtt, 1979; Flora Argentina, 2012; Jardim Botânico do Rio de Janeiro, 2013).
Para la caracterización de la pradera del palmar se estimaron las medias y desvíos de las frecuencias relativas de los taxones para ambas situaciones de muestreo. Se realizó un análisis descriptivo multivariado mediante componentes principales (PCA) y análisis de agrupamiento utilizando distancias euclidianas y el método UPGMA, para lo cual se utilizaron los procedimientos PRINCOMP y CLUSTER del SAS, respectivamente (SAS Institute, 2004).
Se calcularon S (riqueza específica) e índice de diversidad de Shannon H («pi logpi»). Para determinar si existen diferencias significativas entre los estratos CP-SP se utilizaron los siguientes modelos lineales:
Yij = µ + Pi + Tj(i) + eij y Yij = µ + Pi + Aj(i) + eij,
donde:
Yij es S o H, µ es la media general, Pi es el efecto del i-ésimo estrato (CP-SP), Tj(i) es el efecto de la j-ésima transecta larga, Aj(i) es el efecto de la j-ésima transecta corta y eij es el error experimental asociado a la unidad experimental. Tj(i) y Aj(i) son efectos aleatorios.
Para determinar cuáles son los taxones que discriminan las diferencias entre estratos, se utilizó el procedimiento PROC STEPDISC con el método FORWARD y un nivel de significancia del 15 % (Gutiérrez et al., 2003). En este método el programa construye un modelo de discriminación «paso a paso», donde en cada paso realiza un análisis de varianza y selecciona la variable con mayor valor F, introduciéndola en el modelo como covariable; luego procede al siguiente paso.
Recursos fitogenéticos y servicios ecosistémicos
La identificación de los bienes y servicios ambientales, en particular de los recursos fitogenéticos presentes en el área de trabajo, se realizó mediante revisión bibliográfica de la información disponible.
Resultados y discusión
Composición florística
Este trabajo es el primero sobre los campos naturales del palmar de butiá en Uruguay. La diversidad encontrada en el área de estudio (2200 puntos) fue de 70 taxones y 23 familias (Cuadro 1). El número de especies es probablemente superior al número de taxones ya que algunos taxones fueron identificados solo a nivel de género y en un caso a nivel de familia (Cyperaceae). Los valores son del orden de los encontrados para otros tipos de comunidades campestres de Uruguay en áreas de tamaño comparables por Altesor et al. (1999), Texeira y Altesor (2009), Jaurena et al. (2011) y del sur de Brasil (Pillar y Sosinski, 2003; Boldrini, 2006; Muller et al., 2012). El 67,1 % de los taxones fueron identificados por ambas formas de muestreo. Los 12 taxones no identificados en las transectas cortas presentaron frecuencias promedio entre 0,05 y 0,96 % en las transectas largas. Los 11 taxones no identificados en las transectas largas presentaron frecuencias promedio entre 0,02 y 2 % en las cortas.
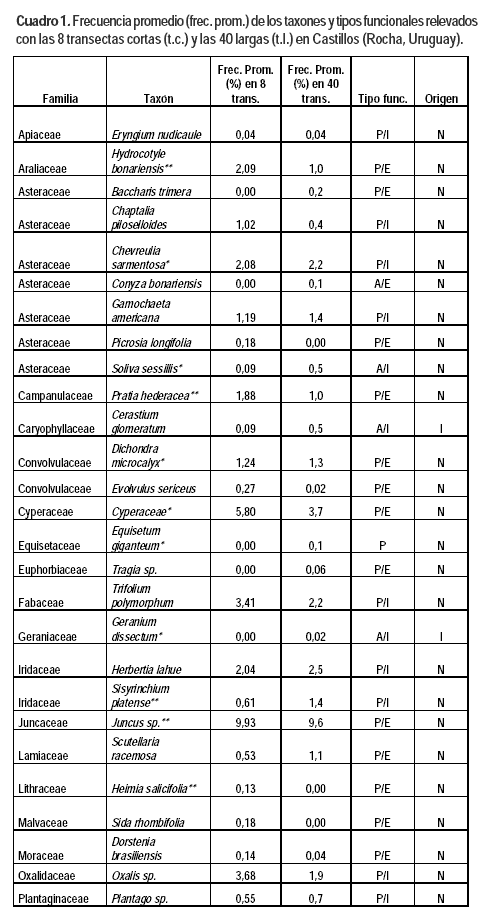
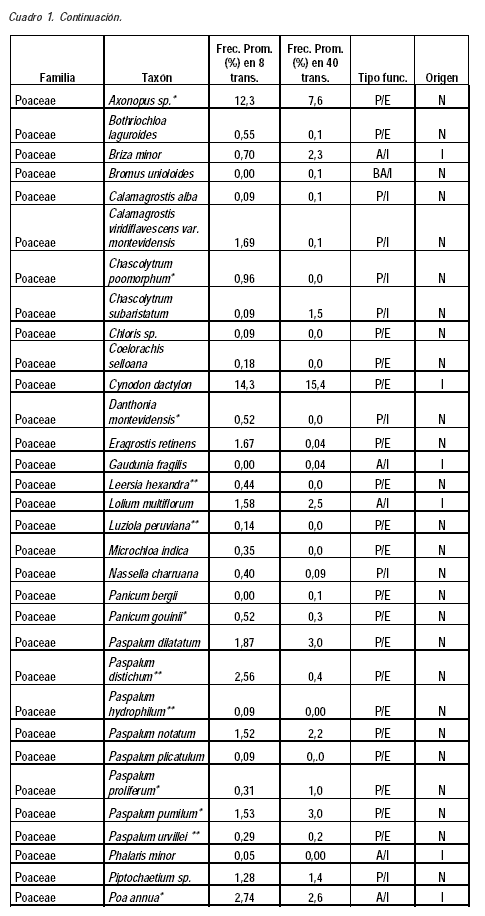
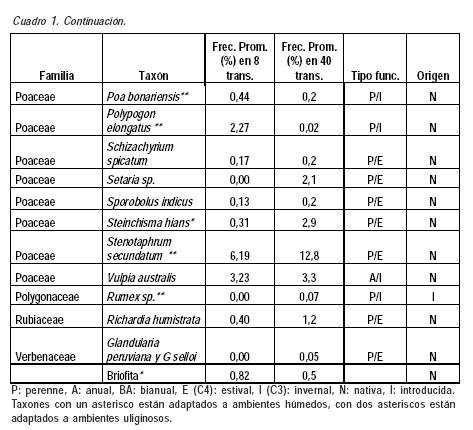
Se comparten familias, géneros y especies considerados característicos del bioma Pampa por Overbeck et al. (2007). El 87,1 % de los taxones presentes son nativos, mientras que la única especie exótica con frecuencia relevante es Cynodon dactylon, y en menor medida Lolium multiflorum. Una situación similar en relación a las exóticas fue registrada por Altesor et al. (1998).
La familia Poaceae representa un 57,1 % de los taxones (82,5 % son perennes), seguido de la familia Asteraceae con un 10 %. Estos datos son concordantes con el hecho que las gramíneas conforman junto con las asteráceas las dos familias más numerosas de los campos de Uruguay y Brasil, seguidos por las leguminosas y ciperáceas (Lezama et al., 2006; Overbeck et al., 2006; Boldrini, 2009).
La presencia de gramíneas C3 (microtérmicas) y C4 (megatérmicas) ha sido señalada como una de las características más destacadas de los campos de la región (Burkart, 1975; Millot et al., 1987; Berretta et al., 2000; Overbeck et al., 2007). En este trabajo la relación de taxones C4/C3 fue de 1,6, lo que demuestra la convivencia de ambos grupos de plantas en los campos de butiá. Cuando se consideran los taxones con frecuencias superiores es notoria la prevalencia de las megatérmicas, situación que se puede explicar tanto porque estos campos presentan suelos húmedos, como por la erosión genética causada por el sobrepastoreo, que afecta principalmente a las gramíneas invernales de hábito cespitoso y provoca la prevalencia de las especies de hábito postrado (Millot et al., 1987; Altesor et al., 1998, 2005; Rodríguez et al., 2003; Overbeck et al., 2007). Entre las especies indicadoras de sobrepastoreo compartidas con otros trabajos se señalan Cynodon dactylon, Richardia humistrata y Chevreulia sarmentosa (Altesor et al., 1998). El pastoreo –principal actividad económica de estos campos– es considerado el principal factor que modela la fisonomía y propiedades ecológicas de los mismos (Pillar y Quadros, 1997).
Diversidad entre campo con palmar y sin palmar
No se detectaron diferencias significativas en los valores de riqueza (S) entre los estratos con y sin palmar, ni cuando se utilizan los datos de las transectas largas (P=0,9305) ni cuando se usan los de las transectas cortas (P=0,3757). Tampoco se detectaron diferencias en diversidad (índice de Shannon) entre estratos por ninguno de los tipos de transectas (para transectas largas: P=0,8473 y para transectas cortas P=0,5209).
Se encontraron diferencias consistentes en la composición florística del estrato herbáceo del palmar en relación al estrato del área circundante sin palmar en todos los casos, independientemente del número y largo de las transectas y del análisis multivariado realizado.
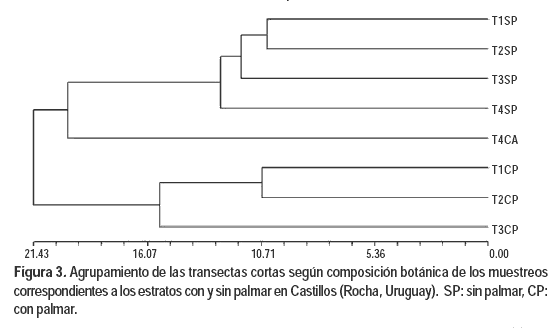
Los agrupamientos de las ocho transectas largas (Figura 3) y de las 40 transectas cortas (Figura 4) muestran que los muestreos correspondientes al campo del palmar conforman un grupo diferenciado de los del campo sin palmas. Los tres primeros grupos que se arman con los datos de las transectas largas acumulan el 84,6% de la variación, correspondiendo los dos primeros grupos (74,9%) a los agrupamientos de dos transectas del estrato con palmar y dos transectas del estrato sin palmar. En el análisis con transectas cortas se alcanza un valor de 85 % con los primeros siete grupos. Estos grupos corresponden en todos los casos a agrupamientos de transectas del mismo estrato, con una predominancia de las transectas del estrato sin palmar. La transecta T4CP (Figura 3) es la primera en agruparse con las transectas del estrato sin palmar debido a que presenta una menor densidad de palmas, dato que corrobora la relación entre composición florística y presencia de palmar.
Los tres primeros componentes del PCA de las transectas largas representan el 26; 19,2 y 15,6 % de la variación total. Mientras que en el análisis con las transectas cortas representan el 14,8; 8,1 y 6,5 % respectivamente. En ambos análisis se observa una nítida separación de las transectas correspondientes al estrato con palmar de las del estrato sin palmar (Figuras 5 y 6), resultado consistente con el análisis de agrupamientos.
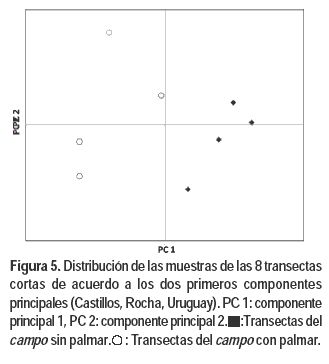
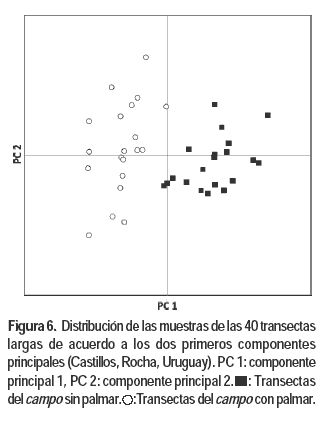
El componente 1 en ambos análisis está explicado básicamente por las diferencias en adaptación de las especies a ambientes húmedos/uliginosos, ya que la topografía y el tipo de suelo es el mismo dentro y fuera del palmar. El sombreado de las palmas reduciría la evapotranspiración y la pérdida de agua de los suelos, situación que estaría generando un microclima de mayor humedad. Los taxones con eigenvalues negativos y superiores a 0,10 se encuentran adaptados a ambientes de suelos más húmedos. Los dos análisis detectaron a los siguientes taxones nativos como característicos del estrato con palmar: Stenotaphrum secuntatum, Hydrocotyle bonariensis, Paspalum urvillei, Juncus spp., Paspalum proliferum y Plantago sp. (Cuadro 1). Otras especies asociadas a los valores negativos del componente 1 y que presentan mayoritariamente adaptación a ambientes húmedos, pero que aparecen sólo en uno de los dos análisis son: Poa bonariensis, Paspalum distichum, Paspalum pumilum, Paspalum hydrophilum, Steinchisma hians, Paspalum dilatatum, Conyza bonariensis y Dichondra microcalyx.
Los taxones más típicos asociados a los valores positivos y superiores a 0,10 de los eigenvalues del componente principal 1 son aquellos que no se encontrarían adaptados a suelos con mayor humedad o que permanecen encharcados durante algunos períodos del año, entre los cuales se encuentran: Calamagrostis montevidensis, Vulpia australis, Chascolytrum subaristatum, Briza minor, Axonopus sp., Paspalum notatum y Setaria sp. El caso de Axonopus sp., característico del estrato sin palmas, se podría explicar por su sustitución funcional por Stenotaphrum secundatum en el tapiz herbáceo del palmar, señalado por su mayor adaptación al sombreado (Burgueño y Nardini, 2009) y a ambientes uliginosos (Rosengurtt, 1979). Por otra parte, si analizamos exclusivamente las especies con frecuencias superiores a 5 %, las especies que separan el estrato palmar del estrato sin palmas son Stenotaphrum secundatum y Juncus spp, ambas asociadas al campo con palmar.
Por otra parte, el resultado del análisis discriminante realizado con el muestreo de las ocho transectas identificó seis especies: Cerastium glomeratum (P=0,0113), Chevreulia sarmentosa (P=0,0273), Coelorachis selloana (P=0,0361), Leersia hexandra (P=0,0077), Pratia hederacea (P=0,0321) y Stenotaphrum secundatum (P=0,0028). En el análisis con los datos de las cuarenta transectas se identificaron 25 especies discriminantes. Ambos análisis identificaron a Stenotaphrum secundatum y Pratia hederacea como características del estrato con palmar, ambas especies citadas como adaptadas a ambientes uliginosos (Rosengurtt, 1979).
El componente 2 del PCA separa transectas en ambos análisis, aunque no se lograron identificar especies características para explicar los resultados. En el análisis con las transectas largas, donde el componente 2 explica una porción razonable de la variación, las cuatro transectas con palmar se separan de acuerdo a la densidad del mismo. Las dos que ocupan el cuadrante superior izquierdo se corresponden a un sector del palmar con baja densidad, mientras que las 2 que ocupan el inferior izquierdo se ubican en un palmar de densidad media a alta. Esto podría estar relacionado con los cambios en exposición a la luz y humedad en ambas situaciones. En el trabajo de Riginos y Grace (2008) la densidad de árboles estuvo asociada con variaciones sustanciales en la riqueza y composición de especies del tapiz herbáceo en la sabana africana. En estudios realizados en el Morro Santana (Porto Alegre, RS, Brasil) se encontró que las parcelas cercanas al borde del bosque diferían claramente en la composición botánica de aquellas que se ubicaban a campo abierto (Overbeck et al., 2006).
Focht y Pillar (2003) reconocieron dos comunidades de campo en Eldorado do Sul (RS, Brasil) de acuerdo al porcentaje de presencia de especies características de cada una de ellas. También encontraron especies con mayor plasticidad ecológica que son las que se encuentran en ambas comunidades y otras que no muestran padrones claros de distribución. Los padrones de vegetación se asociaron a la topografía y al gradiente de humedad de los suelos. Las especies características de las tierras bajas fueron Panicum sabulorum, Paspalum pumilum, Centella asiatica y Eleocharis maculosa.
Lezama et al. (2006) identificó tres unidades principales de vegetación en la región basáltica de Uruguay. Esto estuvo relacionado con un gradiente de disponibilidad de agua, determinado por la profundidad y textura del suelo, y el grado y forma de la pendiente.
En la Pampa húmeda de Argentina, Perelman et al. (2001) reconocieron once tipos de comunidades campestres, fusionadas en cinco unidades de vegetación. En este trabajo se destaca que la principal variación en la composición florística ocurre a pequeña escala, asociada con características topográficas y el gradiente de salinidad de los suelos. La diversidad β, medida de la diferenciación en la composición de especies entre comunidades, indicó mayor heterogeneidad a nivel de paisaje que a nivel regional.
En la sabana Kalakad-Mundanthurai Tiger Reserve (KMTR) de India, Sankaran (2009) encontró alto grado de variación espacial en los padrones de asociación de especies, los que se vinculan con las precipitaciones, el largo de la estación de crecimiento, la presión de pastoreo y la concentración de N, Fe, Mg y K. Los pastos dominantes van cambiando a lo largo del gradiente de elevación, mientras que las especies subordinadas conforman distintos agrupamientos altamente restringidos en su distribución.
De los resultados de este trabajo y de los antecedentes presentados surge que la heterogeneidad de ambientes en los campos y pastizales es un aspecto importante a tener en cuenta para la elaboración de planes de conservación. Reitalu et al. (2012) plantea que esta heterogeneidad produce un incremento de la riqueza de especies especialistas. La conservación de los palmares de B. odorata no sólo debe apostar a la conservación del estrato arbóreo sino también del campo, que presenta características diferenciales desde el punto de vista ecológico y de composición florística.
Bienes y servicios que proveen los campos del palmar de butiá
Mientras que los bosques son considerados remanentes de paisajes prístinos, los biomas campestres han sido considerados hasta tiempos muy recientes como estadios sucesionales secundarios, olvidados por los científicos y sin interés para la conservación. Consecuentemente, la pérdida de la vegetación campestre para la instalación de cultivos y plantaciones forestales ha pasado desapercibida, con escasa oposición (Bond y Parr, 2010).
Behling et al. (2007) sostienen que los campos son ecosistemas naturales que ya existían cuando llegaron los primeros grupos humanos hace unos 12.000 años. Además del forraje para la pecuaria, garantizan la conservación de los recursos hídricos y edáficos, son hábitat de vida silvestre, y ofrecen belleza escénica de potencial turístico (Behling et al., 2009). Los campos son factores importantes de formación de tradición, cultura y economía regionales (Valls et al., 2009).
Como parte de la estrategia global para mantener la biodiversidad en el mundo, existe la necesidad de asegurar que los biomas campestres no sean impactados por los usos antrópicos (Faber-Langendoen y Josse, 2010). Según Heindenreich (2009), el pastoreo extensivo de ganado en los pastizales es considerado la producción de mayor sostenibilidad, uno de los pocos sistemas agrarios compatibles con la conservación de la naturaleza. De los campos se cosechan productos de forma sostenible que contribuyen al bienestar humano y que no destruyen el ecosistema.
Bond y Parr (2010) se refieren a la necesidad de conservar los biomas campestres de especies C4 que han persistido como mosaicos de bosques-pastizales por milenios, principalmente en África, norte de Australia y Sudamérica. La conservación de pastizales y sabanas también es priorizada por Sankaran (2009) fundada en los valores de biodiversidad y en la presencia de plantas de uso medicinal. En este trabajo, el campo se encuentra ubicado en latitud 34 ºS, combinando especies C3 y C4, correspondientes a clima subtropical-templado. El área del palmar podría considerarse como una sabana donde el componente arbóreo son las palmas butiá.
Los campos del palmar de butiá proveen de bienes y servicios a los seres humanos, algunos de valores similares a los que aportan otro tipo de campos del Bioma Pampa, y otros específicos de esta comunidad vegetal. Heindenrich (2009) plantea que los pastizales templados son los ecosistemas más alterados del planeta, con un 41 % de su superficie sustituida por la agricultura, un 13,5 % por otros usos y de lo que resta un porcentaje importante presenta síntomas de degradación. Los estudios sobre los bienes y servicios de estos ecosistemas son prácticamente inexistentes.
En el palmar de butiá los valores de uso directo se vinculan a la pecuaria, la extracción de frutos de butiá tanto para su consumo en fresco como para la elaboración de productos derivados, los recursos fitogenéticos, la producción de miel y la recreación activa (Rivas, 2005). La mayor humedad del suelo y el sombreado ejercido por las palmeras incidirían en la producción de un mayor volumen y calidad de forraje en el período estival. La provisión de sombra y abrigo a los animales son aspectos esenciales para el bienestar animal. El consumo de frutos de butiá por parte del ganado no ha sido evaluado en términos de su incidencia en la nutrición.
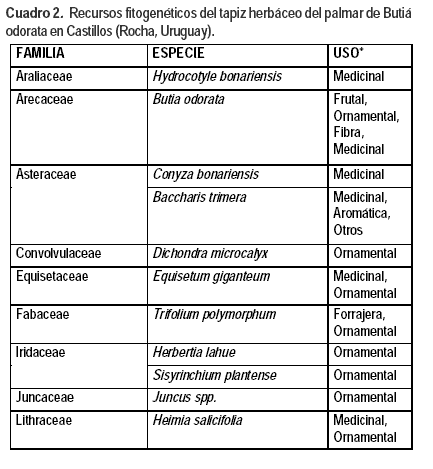
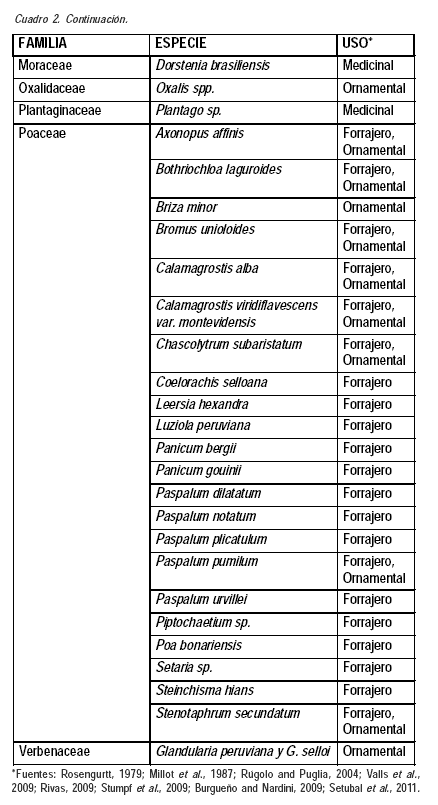
Los recursos fitogenéticos del palmar incluyen especies del tapiz herbáceo, la propia palma butiá y las epífitas que viven sobre las mismas, algunas otras arbóreas presentes en la zona, las especies de gramíneas que forman comunidades de pajonales y las especies de las comunidades de bañados. Este trabajo trata exclusivamente de las herbáceas del campo natural. En el Cuadro 2 se incluye la información obtenida sobre los recursos fitogenéticos presentes en este estudio y los usos citados (Rosengurtt, 1979; Millot et al., 1987; Rúgolo y Puglia, 2004; Valls et al., 2009; Rivas, 2009; Stumpf et al., 2009; Burgueño y Nardini, 2009; Setubal et al., 2011). Los resultados obtenidos indican una alta proporción de recursos fitogenéticos presentes en el palmar, a los que deberían sumarse en un trabajo más exhaustivo las especies de las otras comunidades vegetales que conforman el territorio, caracterizado por una alta diversidad de ambientes (diversidad β).
Sobre los servicios ecosistémicos específicos o diferenciales que brindan los campos del palmar no existen estudios previos, sin embargo se puede señalar que al encontrarse en un ecotono entre los bañados y los campos más altos cumplen un rol en el ciclo hidrológico, en el reciclaje de nutrientes, la conservación de suelos y la regulación del microclima, y son hábitat de vida silvestre.
Los valores socio-culturales del palmar de butiá son ampliamente reconocidos. La belleza escénica, el patrimonio histórico, los conocimientos tradicionales sobre el uso de la palma butiá y las diversas expresiones artísticas confieren a este paisaje y sus habitantes una clara identidad (PROBIDES, 1995; Rivas, 2005; Geymonat y Rocha, 2009).
Los resultados de este trabajo fortalecen los objetivos de conservación del ecosistema palmar de butiá, dando mayor valor al campo natural. El desarrollo de un plan de conservación in situ que contemple tanto la regeneración del palmar como la conservación del campo es una prioridad.
Conclusiones
Este estudio muestra que el campo del palmar de butiá presenta características únicas, donde conviven gramíneas perennes micro y megatérmicas. La composición botánica del estrato herbáceo con palmar es diferente del que no presenta palmas. Las especies que discriminan esta situación son aquellas adaptadas a condiciones de sombreado y suelos húmedos, características del palmar.
El palmar provee de bienes y servicios ecosistémicos, entre los que se destacan la producción de forraje, sombra y abrigo para los animales, los frutos de la palma, la utilización de los recursos fitogenéticos, y la potencialidad que ofrece el ecoturismo.
Agradecimientos
Se agradece al Grupo Palmar y a PROBIDES por el apoyo y colaboración brindada para la ejecución de este trabajo.
Bibliografía
Altesor A, Oesterheld M, Leoni E, Lezama F, Rodríguez C. 2005. Effect of grazing on community structure and productivity of a Uruguayan grassland. Plant Ecology, 179: 83 – 91.
Altesor A, Pezzani F, Grun S, Rodríguez C. 1999. Relationship between spatial strategies and morphological attributes in a Uruguayan grassland : a functional approach. Journal of Vegetation Science, 10: 457-462.
Altesor A, Di Landro E, May H, Ezcurra E. 1998. Long–term species change in a Uruguayan grassland. Journal of Vegetation Science, 9: 173 – 180.
Behling H, Jeske – Pieruschka V, Schuler L, Pillar VD. 2009. Dinâmica dos campos no sul de Brasil durante o Quaternario Tardio. En: Pillar VD, Muller SC, Souza ZM, Avila A. [Eds.]. Campos sulinos : Conservação e uso sustentável da biodiversidade. Brasilia : MMA. pp. 13 - 25.
Behling H, Pillar VD, Muller SC, Overbeck GE. 2007. Late–Holocene fire history in a forest–grassland mosaic in southern Brazil : Implications for conservation. Applied Vegetation Science, 10: 81- 90.
Berretta A, Condon F, Rivas M. 2007. Segundo informe país sobre el estado de los recursos fitogenéticos para la alimentación y la agricultura. Montevideo : MGAP. 120p.
Berretta E, Risso D, Montosi F, Pigurina G. 2000. Campos in Uruguay. En: Lemaire G, Hodgson J, Moraes PC, Carvalho F, Nabinger C. [Eds.]. Grassland ecophysiology and Grazing ecology. Wallingford : CABI Publishing. pp. 377 – 394.
Bilenca D, Miñarro F. 2004. Identificación de Áreas valiosas de pastizal (AVPs) en las Pampas y Campos de Argentina, Uruguay y sur de Brasil. Buenos Aires : Fundación Vida Silvestre. 301p.
Boldrini II. 2009. A flora dos campos do Rio Grande do Sul. En: Pillar VD, Muller SC, Souza ZM, Avila A. [Eds.]. Campos sulinos : Conservação e uso sustentável da biodiversidade. Brasilia : MMA. pp. 63 – 77.
Boldrini II. 2006. Biodiversidade dos campos sulinos. En: Dall´Agnol M, Nabinger C, Rosa LM [Eds.]. Anais do I Simpósio de Forrageiras e Produção Animal. Ênfase: Importância e potencial produtivo da pastagem nativa. Porto Alegre: Departamento de Plantas Forrageiras e Agrometeorologia. UFRGS pp. 11 - 24.
Bond WJ, Parr CL. 2010. Beyond the forest edge : Ecology, diversity and conservation of the grassy biomes. Biological Conservation, 143: 2395 – 2404.
Burgueño G, Nardini C. 2009. Introducción al paisaje natural : diseño de espacios con plantas nativas rioplatenses. Buenos Aires : Orientación Gráfica Editora. 461p.
Burkart A. 1975. Evolution of grasses and grasslands in South America. Taxon, 24: 53 – 66.
Cabrera AL, Willink A. 1973. Biogeografía de América Latina. Washington : OEA. 120p. (Serie de Biología ; 13).
Chebataroff J. 1951. Regiones naturales del Uruguay y de Río Grande del Sur. Revista Uruguaya de Geografía, 2(4): 5 – 40.
Daget P, Poissonet J. 1971. Une method d´analyse phytologique des prairies, criteres d´application. Annales Agronomiques, 22: 5 – 41.
Durán A. 1999. Los suelos del Uruguay. 2a ed. Montevideo : Hemisferio Sur. 358p.
Faber-Langendoen D, Josse C. 2010. World Grasslands and Biodiversity Patterns: A Report to IUCN Ecosystem Management Programme. Arlington: Nature Reserve. 25p.
Flora Argentina. 2012: Plantas vasculares de la República Argentina [En línea]. Consultado 10 noviembre 2012. Disponible en: http://www.floraargentina.edu.ar.
Focht T, Pillar VD. 2003. Spatial patterns and relations with site factors in a campos grassland under grazing. Brazilian Journal of Biology, 63 (3): 423 – 436.
GEO URUGUAY. 2008. Informe del estado del ambiente. Montevideo: CLAES, PNUMA, DINAMA. 350p.
Geymonat G, Rocha N. 2009. M´Botiá : Ecosistema único en el mundo. Castillos: Casa Ambiental. 405p.
Gutiérrez L, Franco J, Crossa J, Abadie T. 2003. Comparing a preliminary racial classification with a numerical classification of the maize landraces of Uruguay. Crop Science, 43: 718 – 727.
Heidenreich B. 2009. What are global temperate grasslands worth? : A case for their protection : A review of current research on their Total Economic Value. Vancouver: Temperate Grassland Conservation Iniciative. 21p.
IBGE. 2012. Mapa de Biomas y de vegetaçao do Brasil [En línea]. Consultado 27 octubre 2012. Disponible en: http://www.ibge.gov.br/home/presidencia/noticias/21052004biomashtml.shtm#sub_download.
Instituto Uruguayo de Meteorología. 1996. Estadísticas climatológicas [En línea]. Consultado 24 octubre 2014. Disponible en: http://meteorologia.gub.uy/ServCli/tablasEstadisticas.
Jardim Botânico do Rio de Janeiro. 2013. Lista de Espécies da Flora do Brasil [En línea]. Consultado 14 octubre 2014. Disponible en: http://floradobrasil.jbrj.gov.br.
Jaurena M, Bentancur O, Ayala W, Rivas M. 2011. Especies indicadoras y estructura de praderas naturales de basalto con cargas contrastantes de ovinos. Agrociencia, 15(1): 103 – 114.
Laterra P, Rivas M. 2005. Bases y herramientas para la conservación in situ y el manejo integrado de los recursos naturales en los campos y pampas del Cono Sur. Agrociencia, 9(1-2): 169 – 178.
Lezama F, Altesor A, Pereira M, Paruelo J.M. 2011. Descripción de la heterogeneidad florística en los pastizales naturales de las principales regiones geomorfológicas de Uruguay. En: Altesor A, Ayala W, Paruelo JM. [Eds.]. Bases ecológicas y tecnológicas para el manejo de pastizales. Montevideo : INIA. (Serie FPTA ; 26). pp. 15 – 32.
Lezama F, Altesor A, León RJ, Paruelo JM. 2006. Heterogeneidad de la vegetación en pastizales naturales de la región basáltica de Uruguay. Ecología Austral, 16: 167 – 182.
Millot JC, Methol R, Risso D. 1987.Relevamiento de pasturas naturales y mejoramientos extensivos en áreas ganaderas del Uruguay. Montevideo : FUCREA-CHPA. 199p.
Muller S, Overbeck G, Pfadenhauer J, Pillar VD. 2012. Woody species patterns at forest–grassland boundaries in southern Brazil. Flora, 207: 586 – 598.
Noblick L. 2011. Validation of the name Butia odorata. Palms, 55: 48 – 49.
Overbeck GE, Muller SC, Fidelis A, Pfadenhauer J, Pillar VD, Blanco CC, Boldrini II, Bothe R, Forneck ED. 2007. Brazil’s neglected biome : The South Brazilian Campos. Perspectives in Plant Ecology, Evolution and Systematics, 9: 101 – 116.
Overbeck GE, Muller SC, Pillar VD, Pfadenhauer J. 2006. Floristic composition, environmental variation and species distribution patterns in burned grassland in southern Brazil. Brazilian Journal of Biology, 66(4): 1073 – 1090.
Paruelo JM, Guerschman JP, Piñeiro G, Jobbágy EG, Verón SR, Baldi G, Baeza S. 2006. Cambios en el uso de la tierra en Argentina y Uruguay : marcos conceptuales para su análisis. Agrociencia, 10(2): 47 – 61.
Perelman SB, León RJC, Oesterheld M. 2001. Cross-scale vegetation patterns of Flooding Pampa grasslands. Journal of Ecology, 89: 562–577.
Pillar VD, Sosinski JR EE. 2003. An improved method for searching functional types by numerical analysis. Journal of Vegetation Science, 14: 323 – 332.
Pillar VD, Quadros FLF. 1997. Grassland-forest boundaries in southern Brazil. Coenoses, 12: 119 -126.
PROBIDES. 1999. Plan Director : Reserva de Biosfera Bañados del Este/Uruguay. Montevideo : Mosca. 159p.
PROBIDES. 1995. El Palmar, la Palma y el Butiá : Ficha Didáctica 4. Montevideo : Productora Editorial. 23p.
Reitalu T, Purschke O, Johansson LJ, Hall K, Sykes MT, Prentice HC. 2012. Responses of grassland species richness to local and landscape factors depend on spatial scale and habitat specialization. Journal of Vegetation Science, 23: 41-51.
Riginos C, Grace JB. 2008. Savanna tree density, herbivores, and the herbaceous community : bottom–up vs. top–down effects. Ecology, 89(3): 2228 – 2238.
Rivas M. 2010. Valorización y conservación de la biodiversidad en Uruguay. En: García Préchac F, Ernst O, Arbeletche P, Pérez Bidegain M, Pritsch C, Ferenczi A, Rivas M. [Eds.]. Intensificación agrícola : oportunidades y amenazas para un país productivo y natural. Montevideo : Universidad de la República. CSIC (Colección Artículo 2). pp. 89 - 109.
Rivas M. 2009. Especies de plantas vasculares de interés socio-económico prioritarias para la conservación. En: Soutullo A, Alonso E, Arrieta D, Beyhaut R, Carreira S, Clavijo C, Cravino J, Delfino L, Fabiano G, Fagúndez C, Haretche F, Marchesi E, Passadore C, Rivas M, Scarabino F, Sosa B, Vidal N. [Eds.]. Especies prioritarias para la conservación en Uruguay : Proyecto Fortalecimiento del proceso de implementación del Sistema Nacional de Áreas Protegidas. Montevideo : GEF, UNDP, FFEM, AECID, DINAMA. (Serie de informes ; 16). pp. 78 -91.
Rivas M. 2005. Desafíos y alternativas para la conservación in situ de los palmares de Butia capitata. Agrociencia, 5(1-2): 161 - 168.
Rodríguez C, Leoni E, Lezama F, Altesor A. 2003. A. Temporal trends in species composition and plant traits in natural grasslands of Uruguay. Journal of Vegetation Science, 14: 433 – 440.
Rosengurtt B. 1979. Tablas de comportamiento de las especies de plantas de campos naturales en el Uruguay. Montevideo : Facultad de Agronomía. Universidad de la República. 86p.
Rosengurtt B. 1943. Estudios sobre praderas naturales del Uruguay : Tercera contribución. Montevideo : Barreiro y Ramos. 281p.
Rúgolo Z, Puglia M. 2004. Gramíneas ornamentales. Buenos Aires : L.O.L.A. 336p.
Sankaran M. 2009. Diversity patterns in savanna grassland communities : implications for conservation strategies in a biodiversity hotspot. Biodiversity Conservation, 18: 1099 – 1115.
Setubal RB, Boldrini II, Ferreria PMA. 2011. Campos dos morros de Porto Alegre. Porto Alegre : Igré, Associação Socio, Ambientalista. 254p.
Soriano A. 1991. Río de la Plata grasslands. En: Coupland RT. [Ed]. Natural grasslands : Introduction and western hemisphere. Amsterdam : Elsevier. pp. 367- 407.
Stumpf ER, Barbieri RL, Heiden G. 2009. Cores e formas no Bioma Pampa : plantas ornamentais nativas. Pelotas : Embrapa Clima Temperado. 273p.
Texeira M, Altesor A. 2009. Small-scale spatial dynamics of vegetation in a grazed Uruguayan grassland. Austral Ecology, 34: 386 – 394.
Valls JFM, Boldrini II, Longhi-Wagner H, Miotto S. 2009. O patrimonio florístico dos Campos : potencialidades de uso e a conservação de seus recursos genéticos. En: Pillar VD, Muller SC, Souza ZM, Avila A. [Eds.]. Campos sulinos: Conservação e uso sustentável da biodiversidade. Brasilia : MMA. pp. 139 – 154.
Zaffaroni C, Rivas M, Resnichenko Y, Hernández J. 2005. Aporte para la conservación de paisajes singulares : el caso de los palmares de Butia capitata (Mart.) Becc., en el departamento de Rocha, Uruguay. En: Anales X Encuentro de Geógrafos de América Latina; Sao Paulo, USP. pp. 16611-16622.

