










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Links relacionados
Compartir

 Permalink
PermalinkAgrociencia (Uruguay)
versión impresa ISSN 1510-0839versión On-line ISSN 2301-1548
Agrociencia Uruguay vol.15 no.1 Montevideo jun. 2011
Especies indicadoras y estructura de praderas naturales de basalto con cargas contrastantes de ovinos
Jaurena Martín1, Bentancur Oscar 2, Ayala Walter3, Rivas Mercedes4
1Instituto Nacional de Investigación Agropecuaria (INIA) Tacuarembó. Ruta 5 km 386, Tacuarembó, Uruguay. Correo electrónico: mjaurena@tb.inia.org.uy
2Departamento de Biometría, Estadística y Computación. Estación Experimental Mario A.Cassinoni (EEMAC), Facultad de Agronomía, Universidad de la República.
3Instituto Nacional de Investigación Agropecuaria, INIA Treinta y Tres.
4Departamento de Biología Vegetal. Facultad de Agronomía, Universidad de la República.
Recibido: 9/3/10 Aceptado: 11/11/10
Resumen
El ajuste de la carga animal es una medida de manejo muy importante en sistemas pastoriles, debido a que determina la relación de la oferta de forraje con la demanda animal, y con ello afecta la productividad animal. Un experimento evaluó el efecto de 5,4 y 10,8 capones ha-1 en la estructura de praderas naturales sobre Litosoles negros y rojos de basalto en Uruguay. Se evaluó la disponibilidad y altura del forraje, composición de especies y diversidad florística y se identificaron especies indicadoras de los niveles de carga animal. El aumento de la carga animal disminuyó la oferta de forraje, pero no se detectaron cambios en la riqueza y diversidad de especies. La variable tipo de suelos explicó la mayoría de las diferencias en la composición botánica, mientras que la respuesta al incremento de la carga animal fue de menor magnitud. La respuesta de la vegetación frente al incremento de la carga de animales fue diferente según el tipo de suelo. En Litosoles negros el incremento de la carga provocó la sustitución de gramíneas perennes cespitosas de ciclo invernal por especies postradas, mientras que en Litosoles rojos este cambio fue menor. En los Litosoles negros, en los niveles de carga animal evaluados existen dos estados alternativos de la vegetación caracterizados por la dominancia de gramíneas cespitosas y postradas. En cambio, en los Litosoles rojos en ambas cargas dominaron especies postradas. Se identificaron 12 especies indicadoras de los niveles de carga animal, las cuales podrían ser utilizadas para evaluar la historia reciente de pastoreo. La evaluación de estas especies facilitará el desarrollo de planes de manejo y monitoreo de praderas naturales de basalto.
Palabras clave: PRADERAS, PASTOREO, OVINOS, DOTACIÓN, VEGETACIÓN
Summary
Indicator Species and Structure of Natural Grasslands on Basaltic Soils with Contrasting Sheep Stocking Rates
The stocking rate adjustment is one of the most important management tools in grasslands ecosystems, since it determines the relationship of forage offer with animal demand, and thereby affects animal production. A field experiment assessed the effect of 5.4 vs 10.8 wether ha-1 on the structure of grasslands of red and black basaltic shallow soils of Uruguay. Forage availability and height, species composition and floristic diversity were evaluated, and indicator species of stocking rate levels were identified. The increase of the stocking rate decreased the offered forage biomass, but no changes were detected in species richness and diversity. The soil type factor explained most of the botanical composition differences, while the response to stocking rates was of smallermagnitude. The vegetation response to the stocking rate increase was different between soil types. For the evaluated stocking rates, Black Lithosols showed two different vegetation states characterized one by the dominance of cool season tall grasses and the other by prostrate species. On the other hand, the botanical composition on the Red Lithosols was little affected. Twelve indicator species related to stocking rates levels were identified, which can be used as indicators of the recent history of grazing. The evaluation of this species will facilitate the development of management and monitoring plans of Basaltic grasslands.
Key words: GRASSLANDS, GRAZING, STOCKING RATE, SHEEP, VEGETATION
Introducción
Los campos o praderas naturales de Uruguay integran el Bioma Campos, que junto al Bioma Pampas forman los pastizales del Río de la Plata, una de las regiones de praderas templadas más extensa del mundo (Soriano, 1991). Estas praderas naturales presentan valores únicos de biodiversidad con más de tres mil especies de plantas vasculares de clima templado y subtropical (Bilenca y Miñarro, 2004), destacándose por la coexistencia de gramíneas C3 y C4. Las praderas naturales son la base nutricional de la ganadería de Uruguay, pero están ocurriendo cambios en el uso de la tierra que amenazan su conservación (Laterra y Rivas, 2005; Díaz, et al., 2006). En este contexto, es necesario desarrollar estrategias destinadas a conservar la diversidad biológica de las mismas.
El principal factor determinante del tipo de vegetación de pradera en Uruguay es el tipo de suelo (Millot et al., 1987; Berretta, 1998; Olmos y Godron, 1990). Los suelos de basalto se originaron a partir de derrames de lava (Bossi y Navarro, 1988) y ocupan 4.1 millones de hectáreas, casi una cuarta parte de la superficie agropecuaria de Uruguay (MGAP, 1979). Los suelos superficiales comprenden dos tercios de la región de Basalto (MGAP, 1979) y tienen aptitud de uso exclusivamente pastoril para la cría de vacunos y ovinos. Las praderas naturales de Basalto han sido agrupadas según los tipos de suelos dominantes en: Litosoles rojos, Litosoles negros, suelos medios y profundos (Castro, 1980; Berretta y Bemhaja, 1998). La heterogeneidad de estas praderas fue clasificada por Lezama (2005) en tres unidades de vegetación y seis comunidades, destacando la disponibilidad de agua como el principal factor de control ambiental que explica los gradientes florísticos y funcionales.
El ajuste de la carga animal es una de las herramientas más importantes en el manejo de praderas naturales (Manley et al., 1997; Holechek et al., 1999), debido a que se modifica la producción individual y por unidad de superficie. Al fijar la carga animal se establece la demanda animal sobre la pradera y se afecta la oferta de forraje. Las respuestas estructurales y funcionales de las praderas naturales al incremento del pastoreo pueden ser muy disímiles y a veces opuestas. Paruelo et al. (2004) atribuyen estas diferencias a la falta de definición en el nivel y escala de análisis de los diferentes trabajos. Estas respuestas disímiles también han sido relacionadas con diferencias en: la historia evolutiva del pastoreo, el tipo de herbívoro que pastorea, el tipo de suelo y su productividad (Milchunas et al., 1988), las condiciones del clima (Fynn y O’Connor, 2000) y el estado inicial de la vegetación (Pickett et al., 1987).
La estructura de las praderas naturales se refiere a la configuración espacial y a la composición de especies de una comunidad (Sala et al., 1986). El pastoreo es un disturbio clave que modifica la estructura y la función de las praderas naturales (Milchunas y Lauenroth, 1993; Rodríguez, et al., 2003; Cingolaniet al., 2005 ; Altesor et al., 2006). Los principales componentes de la estructura de las praderas que son afectados por el pastoreo son la composición y diversidad de especies (Noy-Meir et al., 1989; McIntyre y Lavorel, 2001; Rodríguez et al., 2003) así como la oferta de forraje y su distribución vertical. Existe acuerdo en que incrementos en la intensidad de pastoreo aumentan la tasa de utilización de la pastura y el pisoteo (Borrelli, 2001), aceleran la erosión de suelos y disminuyen la frecuencia de las especies y ecotipos más valiosos de la pradera natural (Millot, et al., 1987). Milchunas, et al. (1988) propusieron un modelo que establece, para sitios con corta historia evolutiva de pastoreo de herbívoros domésticos como las praderas de Uruguay, una rápida disminución de la diversidad y escasa resiliencia frente al aumento de la intensidad de pastoreo. Ante estos posibles efectos negativos del incremento de la presión de pastoreo es necesario monitorear los cambios en el estado de degradación de las praderas naturales. En este sentido, la determinación de las especies indicadoras crecientes y decrecientes frente al incremento de la carga animal constituye una herramienta útil para identificar el estado de conservación de la pradera.
En Uruguay se está promoviendo el desarrollo de sistemas de producción de lanas finas en sistemas pastoriles sobre suelos superficiales de basalto (Montossi et al., 2005). Existen propuestas tecnológicas para incrementar la producción de lana fina basadas en el aumento de la carga de ovinos, ante lo cual surge la necesidad de investigar los impactos de dicha medida en el ecosistema. Esta situación se da en un contexto de creciente preocupación por conocer los efectos de la intensificación productiva en la sostenibilidad de las praderas naturales de Uruguay (Díaz et al., 2006).
La hipótesis del presente trabajo se basa en que el incremento de la carga animal modifica la estructura de praderas naturales. Se espera que el incremento de la carga animal disminuya la biomasa y la altura de la pradera, la frecuencia de gramíneas perennes cespitosas y la diversidad de especies. Para lo cual, se plantean dos objetivos: 1) evaluar el efecto del incremento de la carga de ovinos en la estructura de comunidades vegetales en dos tipos de suelos superficiales de Basalto y 2) identificar especies indicadoras de dos niveles contrastantes de carga animal.
Materiales y métodos
En la Unidad Experimental Glencoe, Paysandú-Uruguay (latitud 32o01´57"S, longitud 57o13´52"W), en el período setiembre 2006 a noviembre 2007 se estudió el efecto de dos cargas de ovinos en la estructura de praderas naturales sobre suelos superficiales de Basalto. Los suelos predominantes fueron Litosoles subéutricos melánicos (Litosoles rojos) y Litosoles éutricos melánicos (Litosoles negros) correspondientes a la unidad Queguay Chico de la carta de reconocimiento de suelos (MGAP, 1979). A fines del invierno 2006 el tratamiento de alta carga se suplementó con fardos en para evitar la muerte de animales.
El diseño experimental fue de parcelas divididas; las parcelas principales se establecieron según el tipo de suelos: 2,2 ha sobre Litosoles rojos y 2,2 ha sobre Litosoles negros. Los Litosoles negros presentan mayores valores de fósforo, carbono orgánico y profundidad que los rojos, de acuerdo con reportes previos de Durán (1985). Cada parcela principal se dividió en dos parcelas menores de 1.1 ha donde se ubicaron al azar los tratamientos de 5,4 capones ha-1 (carga promedio de predios con alta proporción de suelos superficiales) y 10,8 capones ha-1. En cada parcela menor se ajustó la carga de ovinos con seis capones y 12 capones Merino Australiano de 50 a 60 kg de peso vivo. Estas parcelas se subdividieron en dos subparcelas (pseudo-réplicas) de 0,55 ha, ubicadas en la posición topográfica alta y baja de cada tratamiento de carga, en las que se realizó pastoreo alterno cada 21 días. En cada subparcela se establecieron cuatro transectas de 25 metros de longitud sobre suelos superficiales de 10 a 20 centímetros de profundidad. En total se ubicaron 32 transectas en las ocho subparcelas, que fueron las unidades básicas para el seguimiento de la estructura de la vegetación.
Variables evaluadas
Disponibilidad y altura del forraje
En cada subparcela se estimó forraje disponible cada 42 días mediante cortes al ras del suelo con tijera de aro en dos rectángulos de 20 x 50 cm por transecta y se determinó la altura promedio del tapiz tomando ocho registros por transecta. Se registró el peso verde de cada corte y se tomaron submuestras del mismo para la determinación del porcentaje de materia seca en estufa a 70 oC durante 72 horas. Para analizar la variable disponibilidad de forraje se ajustó un modelo lineal general considerando el tipo de suelo, la carga animal y la estación del año como efectos fijos, y las subparcelas de pastoreo alterno (combinación tipo de suelo-carga animal) como efecto aleatorio de acuerdo con el siguiente modelo Yijk = m + Cargai + Sueloj + Estaciónk + Subparcelal + Cargai* Sueloj + Sueloj * Estaciónk + Cargai * Estaciónk + Cargai* Sueloj * Estaciónk + εijkl. Si bien no existieron repeticiones reales, las subparcelas de pastoreo alterno se consideraron como pseudo-repeticiones y la variación entre las subparcelas se utilizó como error experimental, vulnerando el supuesto de independencia de las repeticiones. A pesar de estas condiciones, el cluster de presencias y ausencias de especies demostró que las subparcelas de un mismo tratamiento no fueron más parecidas entre sí que el resto de las mismas, situación que levanta parcialmente la restricción de falta de independencia. Las medias de los efectos significativos fueron comparadas usando el test de Tukey al 5%. Se usó el procedimiento MIXED del paquete estadístico SAS versión 9.1.3 (SAS Institute, 2005). Para la variable altura de forraje se estableció un modelo de regresión lineal con la disponibilidad de forraje.
Composición botánica y diversidad de especies
La composición botánica se evaluó en un sólo momento (noviembre 2007) debido a que los posibles cambios en la composición de especies se esperan en el largo plazo. En dicha evaluación se utilizaron los métodos Point quadrat (Daget y Poissonet, 1971) y Botanal modificado (Millot y Saldanha, 1998). El método Point quadrat se utilizó en cada transecta relevando todas las especies presentes en puntos de contacto cada 50 centímetros. Esta técnica permite estudiar la distribución de las especies mediante el cálculo de frecuencias específicas, índices de riqueza y diversidad (Daget y Poissonet, 1971), El método Botanal se utilizó para registrar la contribución específica (biomasa de una especie/biomasa del total de especies) por apreciación visual con un mínimo de 5%, en 10 cuadros fijos de 0,25 m2 ubicados equidistantemente en cada transecta. Esta técnica es indicada para estudiar las relaciones de dominancia entre las diferentes especies de una comunidad (Tothill et al, 1978).
Con los registros de los censos de composición botánica con el método Point quadrat se generó una base de datos primaria de 102 especies y 32 transectas. En dichos registros se incluyó el porcentaje de suelo desnudo como una especie más, debido a que la falta de especies es un dato de importancia ecológica. Se eliminaron 16 especies de la referida base de datos porque estuvieron presentes en una única transecta y a cuatro transectas «outliers» o fuera de tipo. Estas especies y transectas outliers se eliminaron por presentar más de dos desvíos estándar respecto a la media de las distancias multivariadas según lo propuesto por Mc Cune y Grace (2002), quedando finalmente 86 especies y 28 transectas. Con esta base de datos depurada del método Point quadrat se construyeron dos matrices: i) Presencias y ausencias de especies y ii) Frecuencias específicas. Los datos de presencias y ausencias se agruparon por subparcelas y se realizó un análisis multivariado de aglomeración utilizando el método de Ward como algoritmo de fusión y el índice de Jaccard como medida de distancia. Con la matriz de frecuencias específicas se analizaron los efectos de la carga animal y el tipo de suelo en el número total de especies (riqueza de especies), y en el índice de diversidad de especies Shannon-Wiener (Braun Blanquet, 1979). Para estas variables se ajustó un modelo lineal general similar al utilizado para analizar la variable disponibilidad de forraje (excepto el efecto estación del año, por ser un factor medido solamente en una oportunidad), previa transformación raíz cuadrada de las mismas con el objetivo de normalizarlas.
La base de datos de contribución específica obtenida por el método Botanal modificado consistió en 97 especies considerando el porcentaje de suelo desnudo (columnas) y 32 transectas (filas). Posteriormente se eliminaron 17 especies presentes en una única transecta y cuatro transectas «outliers», quedando una matriz de 80 columnas y 28 filas. Con esta matriz depurada se realizaron análisis de varianza de la contribución porcentual de las principales familias botánicas, ciclos de vida y hábitos de crecimiento con el mismo modelo utilizado para evaluar la diversidad y riqueza de especies. También se realizó un análisis de Componentes Principales para obtener los coeficientes de regresión de la variables suelo y carga animal con los dos primeros componentes. A partir de estas regresiones se determinó el porcentaje de variación que explicó cada una de las variables. Con las 24 especies más abundantes (contribución específica promedio mayor a 0,8%) de la matriz depurada del método Botanal, que en su conjunto acumularon el 75% de la contribución específica, se realizó un análisis multivariado de Correspondencia Canónica con el programa PC-ORD 5.0 (McCune y Mefford, 2006). En dicho análisis se utilizó la distancia de Sorensen, que es una medida del porcentaje de disimilaridad (McCune y Grace, 2002). Este análisis se realizó con el propósito de visualizar las trayectorias de cambio en la composición botánica asociadas al incremento de la cargas de ovinos y al tipo de suelo.
Especies indicadoras
Con los registros de la matriz depurada del método Botanal, en cada tipo de suelo se determinaron las especies indicadoras (crecientes y decrecientes frente al incremento de la carga de capones) utilizando el método INDVAL propuesto por Dufrene y Legendre (1997). Dicho método determina un valor indicador para cada especie y su probabilidad, basado en la especificidad (exclusividad en un hábitat) y en la fidelidad (frecuencia de ocurrencia en el hábitat). Se consideraron como especies indicadoras aquellas que presentaron un valor indicador mayor de 70 y significativo (p<0.05).
Resultados y discusión
Disponibilidad y altura del forraje
La variabilidad climática junto a las diferencias entre los Litosoles provocaron una interacción triple (p < 0,01) de la carga de ovinos con el tipo de suelo y la estación del año en la variable disponibilidad de forraje. En la primavera 2006 no ocurrieron diferencias en la disponibilidad de forraje entre los tipos de suelos, ni entre los niveles de carga animal, probablemente debido a que las precipitaciones se ubicaron debajo de los promedios históricos. En verano 2007 sólo la carga 5,4 capones ha-1 en Litosoles negros presentó mayor disponibilidad de forraje que la carga 10,8 capones ha-1 en Litosoles rojos. En cambio, en otoño e invierno 2007 los Litosoles negros presentaron mayor disponibilidad de forraje que los rojos, mientras que el incremento de la carga animal disminuyó la oferta de forraje sólo en Litosoles negros (Figura 1). Es durante el período de verano y otoño de 2007 que se revierte la situación de sequía, con el consecuente incremento en la disponibilidad de forraje. La altura del forraje presentó una asociación lineal con la disponibilidad de forraje, ajustando el modelo de relación funcional: Disponibilidad de forraje = 229 kg MS ha-1 + 304 kg MS ha-1 por centímetro de altura del forraje (R2 = 0,76, p <0,0001). Durante todo el año 2006 las precipitaciones se ubicaron debajo de los promedios históricos.
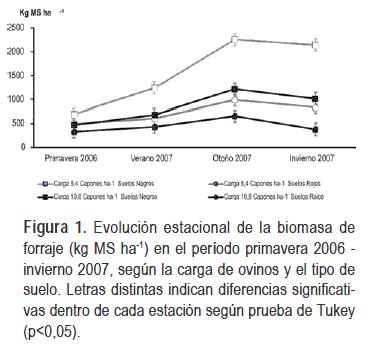
La interacción de la carga con el tipo de suelo en la disponibilidad de forraje es explicada por las diferencias en la composición de especies de las comunidades y por las respuestas heterogéneas que éstas presentan frente a cambios en el clima. La disminución de la biomasa y altura del forraje al incrementar la carga de capones en Litosoles negros se relaciona con una mayor demanda animal que reduce la oferta de forraje de acuerdo a lo propuesto por Borrelli (2001). En los Litosoles rojos, la menor profundidad del suelo y los niveles más bajos de diversidad de especies determinan un mayor impacto negativo de las sequías y consecuentemente un período de recuperación post-sequía más largo. Tilman y Downing (1994) demostraron en un experimento de largo plazo que las comunidades de praderas naturales con mayor diversidad de especies fueron más resistentes y se recuperaron más rápido luego de una sequía. En este sentido Berretta, et al. (2001) reportó un promedio de producción anual de forraje en el período 1980-1994 un 30% mayor en Litosoles negros que en los rojos, aunque dicha diferencia llegó a un 70% en el año 1989 caracterizado por una sequía extrema.
Diversidad de especies
No existió interacción suelo por carga en las variables riqueza y diversidad de especies. Cuando se utilizó el modelo estadístico con dicha interacción, no se encontraron diferencias significativas en riqueza y diversidad entre las combinaciones de suelo y carga. Cuando se excluyó la interacción del modelo, no se encontraron diferencias para carga de capones, pero sí entre los tipos de suelos (Riqueza de especies 59.5 y 47.2, e Índice de diversidad Shannon-Wiener 4.1 y 3.8, para las subparcelas de Litosoles negros y rojos respectivamente). Estos mayores niveles de diversidad de especies en los Litosoles negros comparados con los rojos se relacionarían con que comunidades sobre suelos más productivos pueden mantener un mayor número de especies. Estos resultados están de acuerdo con Stohlgren et al. (1999) que reportaron un mayor efecto del tipo de suelos y del clima en la diversidad de especies comparado con el pastoreo. La falta de cambios en la riqueza y diversidad de especies frente al incremento de la carga animal no concuerda con la clasificación de corta historia evolutiva de pastoreo propuesta para este tipo de praderas en el modelo de efectos del pastoreo de Milchunas et al. (1988).
Si bien las praderas de Uruguay presentan una corta historia de pastoreo por herbívoros domésticos, en los suelos superficiales de Basalto, habrían existido niveles importantes de presión de pastoreo de herbívoros nativos que explicarían la alta resiliencia de estas comunidades. En este sentido Behling et al. (2009) señalan que los grandes herbívoros extintos, con hábitos de pastoreo similares al de los bovinos, habrían sido responsables del mantenimiento de la alta diversidad de especies en praderas de Río Grande do Sul similares a las de Uruguay. La ocurrencia de selección natural convergente por adaptación a la sequía y a altas intensidades de pastoreo planteada por Milchunas et al. (1988) y Cingolani et al. (2005) también explicaría la alta resiliencia a cambios en la intensidad del pastoreo de estas comunidades sobre suelos superficiales.
Composición de especies
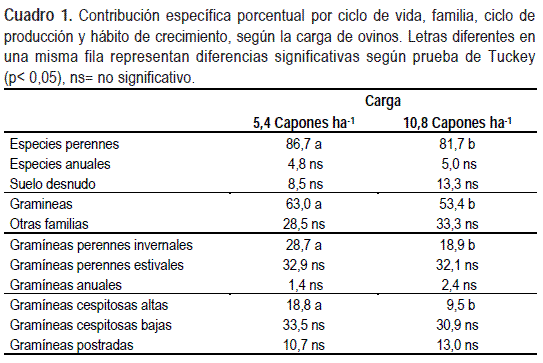
No existieron interacciones significativas suelo por carga en las variables de composición de especies agrupadas por ciclo de vida, familia botánica, ciclo de producción y hábito de crecimiento. Cuando se excluyó la interacción del modelo no ocurrieron diferencias entre los tipos de suelos pero sí entre las cargas de capones, por lo cual se presentan sólo los datos de las cargas de animales. El incremento de la carga de ovinos disminuyó 9,6% la contribución específica de la familia Poaceae (Gramíneas) y 5% de especies perennes en general, mientras que no se detectaron diferencias significativas en otras familias, especies anuales, ni en la proporción de suelo desnudo. Dentro de las gramíneas, el incremento de la carga de ovinos disminuyó la contribución específica de especies perennes invernales un 9,8%, cambio que fue explicado principalmente por una reducción del 9,3% en la contribución de especies cespitosas altas, mientras que no se detectaron diferencias en gramíneas perennes estivales, cespitosas bajas y postradas (Cuadro 1). La disminución de las gramíneas ha sido reportada como uno de los efectos principales del incremento de la intensidad de pastoreo (Lavorel, et al., 1997; McIntyre y Lavorel, 2001). El hábito de crecimiento erecto de algunas especies de gramíneas es destacado por Milchunas et al. (1988) como una adaptación que les permite tener alta capacidad de competencia por luz, pero hace que sean muy vulnerables al pastoreo. En este sentido McIntyre y Lavorel (2001) señalan a las gramíneas cespitosas palatables como el grupo más sensible al aumento de la carga animal. Estas especies no pueden mantener la tasa de crecimiento con el aumento de la intensidad de pastoreo y quedan en desventaja frente a las especies postradas. En este contexto, Millot et al. (1987) y Formoso (1995) señalan que el incremento de la carga de animales por encima de la capacidad de carga de las praderas produce un cambio en la vegetación hacia comunidades dominadas por especies menos productivas.
El análisis multivariado de aglomeración de la matriz de presencias y ausencias de especies relevadas con el método Point quadrat generó un dendograma que clasificó las subparcelas según tipo de suelo, pero no se constató relación con la carga de ovinos (Figura 2). En este análisis no se visualizó una mayor asociación de las subparcelas de un mismo tratamiento de carga animal, atenuando los posibles efectos negativos de la falta de independencia de las pseudoréplicas.
La regresión de los dos primeros componentes principales de la contribución específica, relevada con el método Botanal, con las variables tipo de suelo y carga animal confirmó nuevamente que el tipo de suelo explicó una mayor proporción de la variación en la contribución específica (11,2%) comparado con la carga de ovinos (2,1%). Este mayor efecto del tipo de suelo en la variación florística, se explica por las diferencias en la composición de especies de los Litosoles y confirmaría la resiliencia de estas comunidades al incremento de la intensidad del pastoreo.
El ordenamiento multivariado de los censos florísticos por correspondencia canónica de las 24 especies más abundantes relevadas con el método Botanal explicó el 45,7% de la variación en los dos primeros ejes. El primer eje acumuló el 31,3% de la variación y se relacionó con el tipo de suelo, mientras que el segundo eje explicó sólo el 14,4% y se asoció con la carga de ovinos, aunque se detectó un efecto diferencial del incremento de la carga según el tipo de suelo. En Litosoles negros se pudo establecer una trayectoria de cambio en la composición de especies relacionada con las cargas de animales, mientras que en los Litosoles rojos no se diferenció claramente. En la Figura 3 se presenta un biplot, y un gráfico conjunto de variables, subparcelas y especies, que es una aproximación de la distribución multivariada reducida a un espacio normalmente de dimensión de dos ejes (Gabriel, 1971). En este gráfico se presenta el patrón de variación de la composición de especies de las subparcelas relacionadas con las cargas de ovinos y los tipos de suelos.
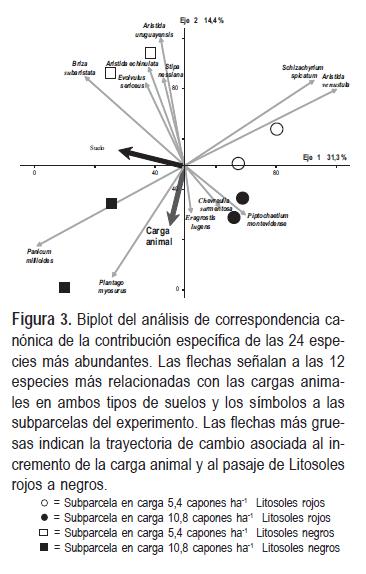
Las especies más relacionadas con la carga 5,4 capones ha-1 en Litosoles negros fueron las gramíneas cespitosas Aristida echinulata Roseng. et Izag., Aristida uruguayensis Henrard, Briza subaristata Lam., Stipa nessiana Trin. y Rupr. y la hierba enana palatable Evolvulus sericeus Sw., mientras que la gramínea cespitosa baja Panicum milioides Nees ex Trin. y la hierba enana anual Plantago myosurus Lam. fueron las más relacionadas con la carga 10,8 capones ha-1. Las especies más relacionadas con la carga 5,4 capones ha-1 en Litosoles rojos fueron las gramíneas cespitosas Aristida venustula Arechav., y Schizachyrium spicatum (Spreng.) Herter, mientras que presentaron niveles intermedios de relación con la carga 10,8 capones ha-1 la hierba enana Chevreulia sarmentosa (Pers.) S.F.Blake y las gramíneas cespitosas bajas Piptochaetium montevidense (Spreng.) Parodi y Eragrostis lugens Nees.
La sequía ocurrida al comienzo del período experimental habría frenado los cambios direccionales provocados por el incremento de la intensidad de pastoreo, explicando así las escasas modificaciones en la composición de las praderas asociados a los niveles de carga animal. Si bien, la información existente para las praderas naturales de Uruguay señala que los principales cambios provocados por la exclusión de herbívoros domésticos ocurren en los dos años siguientes a la clausura del pastoreo (Rodríguez, et al., 2003; Jaurena y Rivas, 2005), los cambios direccionales provocados por cargas de animales contrastantes sucederían en escalas temporales mayores a la considerada en este experimento. En los Litosoles negros, en los niveles de carga animal evaluados existen estados alternativos de la vegetación caracterizados por la presencia de gramíneas cespitosas y postradas. Dichos estados de la vegetación podrían ser reversibles o no en función de la intensidad de pastoreo (definida por la carga animal y por las condiciones climáticas que determinan el crecimiento de la pradera). En cambio, en los Litosoles rojos prácticamente no ocurren cambios en el estado de la vegetación al pasar de 5,4 ovinos ha-1 (carga corrientemente utilizada en predios ovejeros con alta proporción de suelos superficiales) a 10,8 ovinos ha-1. Esta información confirma que en potreros dominados por Litosoles rojos es necesario manejar cargas menores a las utilizadas actualmente, o aplicar alivios estratégicos del pastoreo para permitir la recuperación de las gramíneas perennes cespitosas (Berretta et al., 2001). Al disminuir la intensidad de pastoreo, se podría revertir el sobrepastoreo y permitir que especies clave recuperen la biomasa y aumenten su frecuencia, modificando el estado de la pradera hacia uno caracterizado por gramíneas cespitosas.
Especies indicadoras
Se encontraron nueve especies indicadoras de las cargas de ovinos en Litosoles negros y cuatro especies en los rojos, de las cuales una es P. myosurus de ciclo de vida anual y el resto son perennes (Cuadro 2), confirmando nuevamente que los cambios florísticos fueron más acentuados en Litosoles negros que en los rojos. Estos resultados están de acuerdo con Milchunas y Lauenroth (1993) que reportaron un mayor cambio en la composición de especies como respuesta al aumento de la intensidad de pastoreo en sitios más productivos. Los Litosoles rojos, de menor productividad que los negros (Berretta, et al., 2001) han sobrellevado históricamente mayores presiones de pastoreo, debido a su menor oferta de forraje frente a niveles similares carga animal. En estos suelos la adaptación evolutiva a mayores presiones de herbivoría y la menor fertilidad explicarían la menor respuesta de las comunidades a cambios en la intensidad de pastoreo.
En Litosoles negros, las especies decrecientes con el incremento de la carga de ovinos fueron cuatro gramíneas de hábito cespitoso (A. echinulata, A. uruguayensis, S. spicatum y S. nessiana) y una hierba enana palatable (E. sericeus), mientras que las especies crecientes fueron dos gramíneas estivales, una de hábito postrado y una cespitosa bajo porte (Axonopus affinis Chase y P. milioides Nees ex Trin.) y dos hierbas enanas de hábito muy postrado (P. myosurus y Richardia stellaris (Cham. y Schltdl.) Steud., esta última de muy baja palatabilidad. En Litosoles rojos: las especies decrecientes con el incremento de la carga animal fueron tres gramíneas de hábito cespitoso (A. venustula, Paspalum plicatulum Michx y nuevamente S. spicatum) y como creciente se detectó sólo una gramínea cespitosa de bajo porte (Eragrostis neesii Trin.).
Al utilizar el método INDVAL se detectaron ocho de las 12 especies que ya habían sido relacionadas con las cargas animales en el análisis de correspondencia canónica, logrando resultados similares con ambas técnicas. Todas las especies de gramíneas perennes identificadas como decrecientes poseen hábito de crecimiento cespitoso, en cambio las crecientes tienen hábito de crecimiento más postrado, aunque éstas últimas presentan diferentes mecanismos de respuesta al pastoreo. Cingolani, et al. (2008) señalan que pueden coexistir dos conjuntos de especies crecientes frente a aumentos en la intensidad de pastoreo: uno con especies palatables de crecimiento rápido adaptadas a competir por los recursos (Tolerantes) y otro más adaptado a evitar la herbivoría (Resistentes). Ejemplos de ello son el comportamiento de A. affinis y P. milioides, especies que tolerarían el incremento de la intensidad de pastoreo manteniendo altas tasas de crecimiento en los Litosoles negros. La respuesta creciente de A. affinis estaría explicada por su hábito de crecimiento postrado y por su capacidad de propagación vegetativa, mientras que en P. milioides se relacionaría con su alta capacidad de propagación por semillas. En ambas especies la respuesta creciente también estaría explicada por su alta capacidad de crecimiento en condiciones favorables de humedad de suelos.
Los antecedentes de respuesta al pastoreo de P. milioides son discordantes. Por un lado Rosengurtt (1943) define esta especie como resistente a altas cargas de corta duración, mientras que Altesor, et al. (2005) la clasifican como sucesionalmente intermedia debido al aumento de su frecuencia en los primeros años de exclusión del pastoreo. Este tipo de respuestas contradictorias frente al pastoreo fue reportado por Vesk y Westoby (2001) para el 25% de las especies identificadas como crecientes o decrecientes, lo cual limita la capacidad predictiva de las especies indicadoras. Las respuestas contradictorias estarían relacionadas principalmente con las interacciones de la intensidad y método de pastoreo con el clima, las diferencias genéticas de las poblaciones y la plasticidad fenotípica.
En cambio, P. myosurus, R. stellaris, dos hierbas pequeñas con hábito de roseta, y E. neesii, son especies que minimizan la probabilidad de pastoreo con un hábito todavía más postrado y con hojas poco apetecidas en el caso de R. stellaris. El comportamiento decreciente de la hierba enana palatable E. sericeus se relacionaría con la selectividad de los ovinos sobre la misma.
Conclusiones
El incremento de la carga animal modificó la estructura de las praderas naturales evaluadas, disminuyendo la biomasa aérea, la altura del forraje y la proporción de gramíneas perennes invernales de hábito cespitoso, mientras que no se detectaron cambios en la riqueza y diversidad de especies.
La variable tipo de suelos explicó la mayoría de las diferencias en la composición botánica, mientras que la respuesta al incremento de la carga animal fue de menor magnitud. En las comunidades de Litosoles negros existen dos estados de la vegetación alternativos entre sí, asociados a intensidades contrastantes de pastoreo, caracterizados por la dominancia de gramíneas cespitosas y postradas respectivamente. En cambio en Litosoles rojos se tendrían que manejar cargas menores a 5,4 ovinos ha-1 para llegar al estado caracterizado por gramíneas cespitosas.
Se generó una lista de especies indicadoras de niveles contrastantes de carga animal que pueden ser utilizadas para simplificar la evaluación y monitoreo de la historia reciente de pastoreo, aunque previamente se deberá definir claramente el tipo y estado de las comunidades.
Los resultados del presente estudio deben relativizarse a un año con condiciones climáticas muy particulares, muy secas al inicio y lluviosas al final. A partir de estas evaluaciones recomendamos continuar investigando en experimentos a largo plazo los efectos del incremento de la carga animal en la vegetación y su interacción con el clima.
Agradecimientos
A todos los compañeros de INIA que colaboraron con los muestreos en Glencoe, y a María Behmaja y Laura Silva por sus sugerencias para la corrección del manuscrito. Al Programa de Posgrado en Ciencias Agrarias de la Facultad de Agronomía de la Universidad de la República (Uruguay).
Bibliografía
Altesor A., Piñeiro G. Lezama F., Jackson R., Sarasola M. y Paruelo J.M. 2006. Ecosystem Changes Associated with Grazing in Subhumid South American Grasslands. Journal of Vegetation Science, 17(3): 323–332.
Durán A. 1985. Los suelos del Uruguay. Montevideo: Hemisferio Sur. 398 p.
Milchunas D. and Lauenroth W. 1993. A Quantitative Assessment of the Effects of Grazing on Vegetation and Soils over a Global Range of Environments. Ecological Monographs, 63(4): 327–366.

