










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
Related links
Share

 Permalink
PermalinkAgrociencia (Uruguay)
Print version ISSN 1510-0839On-line version ISSN 2301-1548
Agrociencia Uruguay vol.15 no.1 Montevideo June 2011
Control de la autopolinización, germinación del polen y crecimiento del tubo polínico en mandarina ‘Montenegrina’
Chouza Ximena1, Gravina Alfredo1, Borges Alejandra2
1Departamento de Producción Vegetal. Ecofisiología de Citrus, Facultad de Agronomía, Universidad de la República, Garzón 780, CP 12900. Montevideo, Uruguay. Correo electrónico: xchouza1@gmail.com
2 Departamento de Biometría, Estadística y Computación. Facultad de Agronomía, Garzón 780, CP 12900. Universidad de la República.
Recibido: 13/8/10 Aceptado: 9/11/10
Resumen
El objetivo de este trabajo fue determinar el efecto de distintas combinaciones, concentraciones y número de aplicaciones de GA3, CuSO4 y ANA en la germinación de polen in vitro, crecimiento del tubo polínico en flores tratadas individualmente y la inducción de partenocarpia en campo de la mandarina autocompatible ‘Montenegrina’. El polen se colectó y sembró en medio de cultivo, conformándose nueve tratamientos de distintas dosis y combinaciones de los tres productos. Se seleccionaron botones florales previo a antesis y se les aplicó ANA (200 mg l-1), GA3 (100 mg l-1), GA3 + CuSO4 (100 mg l-1 c/u), CuSO4 (100 mg l-1) y control sin tratar. A los 0, 5 y 10 días post-aplicación se colectaron las flores y se cuantificó en microscopio con fluorescencia el crecimiento del tubo polínico. En campo se seleccionaron 30 árboles y se les aplicaron cinco tratamientos: una y dos aplicaciones de CuSO4 (100 mg l-1), una y dos de GA3 (100 mg l-1) y control. Todos los productos aplicados en el medio de cultivo inhibieron la germinación de polen in vitro y en flores individuales, el crecimiento del tubo polínico con respecto al control. A los 10 días de realizadas las aplicaciones, en ninguno de los tratamientos el avance del tubo polínico superó el 30% de la longitud del estilo y el GA3 (100 mg l-1) impidió su avance más allá del estigma. De los tratamientos a planta entera, el único que resultó eficiente fue la doble aplicación de GA3 que redujo de nueve a siete el número de semillas por fruto en relación al control.
Palabras clave: CuSO4, GA3, SEMILLAS, TUBO POLÍNICO
Summary
Parthenocarpy induction, pollen germination and pollen tube growth of ‘Montenegrina’ mandarin
The objective of this work was to determine the effect of different combinations and concentrations and number of applications of GA3, CuSO4 and NAA in pollen germination in vitro, tube growth in flowers treated individually, and field parthenocarpy induction of self compatible ‘Montenegrina’ mandarin. Pollen was collected and added to culture medium with nine treatments composed by different doses and combinations of the three products. Flower buds previous to anthesis were selected and treated with NAA (200 mg l-1), GA3 (100 mg l-1), GA3 + CuSO4 (100 mg l-1 c/u), CuSO4 (100 mg l-1) and untreated control. At 0, 5 and 10 days post-application the flower buds were collected, and pollen tube growth was measured with fluorescence microscope. Under field conditions 30 trees were selected and five treatments were applied: CuSO4 (100 mg l-1) one and two applications, GA3 100 mg l-1) one and two applications and control. All treatments applied to the culture medium inhibited in vitro pollen germination and tube growth in individual flowers compared to the control. Ten days after application pollen tube grew no further than 30% of the style in any of the treatments and GA3 (100 mg l-1) limited its growth in the stigmatic area. Among the treatments, the only one that was efficient treatment was GA3 double applications, which reduced from nine to seven the number of seeds per fruit compared to the control.
Key words: CuSO4, GA3, POLLEN TUBE, SEEDS
Introducción
La producción de citrus en Uruguay tiene por objetivo la exportación como fruta para consumo en fresco (DIEA, 2008). En los últimos años los consumidores de fruta cítrica fresca han puesto énfasis en la ausencia de semillas como un requerimiento de calidad de fruta (Barry, 2004).
‘Montenegrina’ (Citrus deliciosa), es una de las variedades de mandarina requeridas por los mercados destino de exportación, alcanzando altos precios debido a que presenta muy buenas características organolépticas. Se caracteriza por presentar alta calidad de fruta, recolección tardía y un número menor de semillas por fruto que la original (Anderson, 1996, Rodríguez et al., 1999, Koehler-Santos et al. 2003), de una a 34 y siete promedio para nuestro país (Gambetta et al., 2008). Las flores son completas y autocompatibles, de tamaño pequeño a mediano, desarrollándose preferentemente en brotes uniflorales con o sin hojas. Presenta microesporogénesis regular y elevados índices de fertilidad masculina; el porcentaje de polen viable supera el 95% (Rodrígues y Cunha Dornelles, 1999).
La inducción de partenocarpia, ha sido reportada como una práctica de potencial valor comercial en diferentes cultivos, ya que permite eliminar o disminuir la presencia de semillas sin disminuir la productividad y calidad de frutos.
En citrus se han obtenido resultados positivos en la inducción de partenocarpia con el uso del sulfato de cobre (CuSO4). Las aplicaciones en plena floración en tangor ‘Afourer’ bajo condiciones de polinización cruzada, disminuyeron significativamente el número promedio de semillas por fruto y aumentaron en forma significativa el porcentaje de frutos sin semillas sin reducir el rendimiento. El modo de acción del CuSO4 se ha relacionado con la inhibición de la germinación del polen y la detención del desarrollo del tubo polínico (Mesejo et al., 2006).
Existe evidencia del efecto del ácido giberélico (GA3) aplicado durante el desarrollo de la flor, como inhibidor del desarrollo de semillas en frutos de ‘Ponkan’, ‘Murcott’, ‘Chandler’ (Tominaga, 1997; Domínguez y Rodrígues, 2007; Erner et al., 2008). El modo de acción del GA3 como inductor de partenocarpia en citrus ha sido relacionado con la desincronización del desarrollo masculino y femenino de la flor (Mesejo et al., 2008), disminuyendo el período de polinización efectivo.
La aplicación de auxinas también ha tenido efecto en la reducción del número de semillas. Pulverizaciones de ácido naftalenacético (ANA) a planta entera, tanto en inicio de floración como en frutitos recién cuajados, provocaron una reducción significativa del número de semillas en las mandarinas ‘Dancy’, ‘Murcott’ y ‘Temple’, en tangor ‘Ortanique’ y en pummelo ‘Chandler’ (Feinstein et al., 1975; Lewin y Monselise, 1976; Domínguez y Rodrígues, 2007; Erner et al., 2008). Sin embargo, el modo de acción de las auxinas no está todavía dilucidado.
El objetivo del presente trabajo fue estudiar el efecto de distintas dosis, combinaciones y número de aplicaciones de reguladores de crecimiento (GA3 y ANA) y CuSO4 sobre la germinación de polen in vitro, el crecimiento del tubo polínico en flores tratadas individualmente y la inducción de partenocarpia en condiciones de campo en mandarina ‘Montenegrina’.
Materiales y métodos
Germinación de polen in vitro
Se seleccionaron dos árboles de un cuadro de mandarina ‘Montenegrina’ (Citrus deliciosa Tenore), injertado sobre trifolia (Poncirus trifoliata L.Raf.). En plena floración se recolectaron 30-40 flores iniciando antesis (estado 61 de la escala BBCH) (Agustí et al., 1995). Las flores se colocaron en una caja de petri abierta que se depositó en una cámara con sílica gel durante 24-36 horas para que abrieran las anteras. Luego se mantuvieron las anteras abiertas en heladera durante dos horas para que se hidrataran los granos de polen.
La germinación de los granos de polen se realizó en portaobjetos, colocando una base de 750 µl de medio sólido. De las anteras se extrajeron los granos de polen frotando las mismas con pincel fino para recoger la mayor cantidad de granos de polen, los cuales se depositaron sobre el medio sólido de cada portaobjeto. Posteriormente se cubrieron los granos de polen con 30 µl de medio líquido.
Se utilizó el medio de cultivo Brewbaker y Kwack (1963) (10% sucrosa, 100 ppm H3BO3, 300 ppm Ca (NO3)2 4H2O, 200 ppm MgSO4 y 100 ppm KNO3 con PH 5,4 ± 0,1) y se dividió en dos partes: una se utilizó como medio líquido y la otra como medio sólido con el agregado de Phytagel (polvo incoloro espesante). A su vez ambos medios de cultivo se dividieron en nueve soluciones cada uno.
A cada una de esas soluciones se le agregaron los productos en las concentraciones determinadas para cada uno de los tratamientos. Se utilizó un diseño completo al azar con nueve tratamientos y tres repeticiones (portaobjetos). Los tratamientos fueron:
(1) GA3 (100 mg l-1)
(2) GA3 (200 mg l-1)
(3) ANA (200 mg l-1)
(4) ANA (300 mg l-1)
(5) CuSO4 (50 mg l-1)
(6) CuSO4 (100 mg l-1)
(7) CuSO4 (100 mg l-1) + GA3 (100 mg l-1 )
(8) CuSO4 (100 mg l-1)+ ANA (200 mg l-1)
(9) Control
Se colocaron los preparados en cámara oscura a 25 ºC y 70-80% de humedad hasta que el tratamiento control germinó (a las 72 horas). Una vez que germinaron los granos de polen se fijaron con solución de FAA (formaldehído, acético, etanol al 70%; 5:5:90). La germinación de polen se evaluó contabilizando un promedio de 300-400 granos de polen por repetición con ayuda de un microscopio óptico (OLYMPUS ECE-Bi). Del total de granos contabilizados se determinó el número de granos de polen germinados para calcular el porcentaje de germinación, tomando como criterio que un grano de polen germinó cuando el largo del tubo polínico superaba el diámetro del mismo (Stanley y Linskens, 1974).
Germinación de polen y crecimiento de tubo polínico in vivo
Se seleccionaron dos árboles del mismo cuadro de mandarina ‘Montenegrina’ que se utilizó para el estudio de germinación de polen. Durante la brotación, se eligieron 75 brotes terminales con botones florales alargados, estado 59 de la escala BBCH (Agustí et al., 1995) a los que se le aplicó en grupos de 15 flores los siguientes tratamientos:
1) ANA (200 mg l-1)
2) GA3 (100 mg l-1)
3) GA3 + CuSO4 (100 mg l-1 c/u)
4) CuSO4 (100 mg l-1)
5) Control
La solución de GA3, se acidificó a pH 4,0-4,5 y en todos los casos a las soluciones se les agregó un agente humectante no iónico al 0,05%. Las aplicaciones se realizaron con pulverizadora manual en forma localizada. En el caso del tratamiento combinado de GA3 y CuSO4, se aplicaron por separado sobre las mismas flores ambos productos.
Luego de la aplicación se recolectaron cinco flores por tratamiento en tres momentos: 0, 5 y 10 días post-aplicación. Se fijaron en solución de FAA (formaldehído, acético, etanol 70%; 5:5:90) y se guardaron a 4 ºC hasta que fueron procesadas. Para el análisis se eliminaron los pétalos de las flores, se lavaron con agua destilada y se mantuvieron en sulfito sódico 5%, durante una noche. Al día siguiente, utilizando una nueva solución de la misma concentración de sulfito sódico, los tejidos se ablandaron durante 30 a 40 segundos a 100 watts, en microondas (JAMES-20NDG). Para observar la germinación de los granos de polen y el desarrollo del tubo polínico, se separó el estilo con el estigma del ovario y se seccionó longitudinalmente bajo lupa binocular (OLYMPUS SZ40) con ayuda de bisturí y pinzas. Los pistilos seccionados se colocaron en solución de azul de anilina al 0,1% en K2HPO4 0,1 N (Kearns y Inouye, 1993) donde se mantuvieron por 20 a 30 minutos para teñir los tejidos. Los pistilos seccionados se colocaron en portaobjetos con una gotita de glicerina líquida, se cubrieron con cubreobjetos, evitando la formación de burbujas de aire y se realizó un squash. La evaluación de la germinación de los granos de polen se realizó contando al azar 200-300 granos de polen por flor en tres campos visuales. El desarrollo del tubo polínico en el estilo se determinó como el porcentaje de estilo recorrido por el tubo polínico más largo, utilizando microscopio de fluorescencia Olympus Vanox AH3 con filtro U BH2-DMU para el rango de línea de emisión Hg 334-365 nm. Se utilizó un diseño completo al azar con cinco tratamientos y cinco repeticiones.
Inducción de partenocarpia en condiciones de campo
Al año siguiente de los experimentos anteriores, durante la brotación, se seleccionó un cuadro de la misma variedad en la misma plantación y se seleccionaron 30 árboles que presentaran alta floración, distribuida de forma homogénea, sin deficiencias nutricionales evidentes y en buen estado sanitario. Se utilizó un diseño de bloques completos al azar con un árbol como unidad experimental y seis repeticiones por tratamiento. Se utilizaron las filas del cuadro elegido como bloques en base a que presentaron un manejo similar. Los tratamientos aplicados fueron:
(1) GA3 100 mg l-1 única aplicación 30-40% de flor abierta
(2) GA3 100 mg l-1 doble aplicación 30-40% y 70-80% de flor abierta
(3) CuSO4 100 mg l-1 única aplicación 30-40% de flor abierta
(4) CuSO4 100 mg l-1 doble aplicación 30-40% y 70-80% de flor abierta
(5) Control
Las aplicaciones se realizaron con pulverizadora de puntero a una presión de 10 - 12 bar, cubriendo todo el follaje hasta punto de goteo. La solución de GA3 se ajustó a un pH de 4,0-4,5, a la cual se agregó un humectante no iónico al 0.05%. En el caso del CuSO4 se le agregó el mismo agente humectante a igual dosis. El gasto promedio por planta de ambas soluciones de productos aplicados fue de 7,3 l y 6,7 l. Luego del final de caída fisiológica se realizó raleo de frutos en forma manual utilizando el criterio comercial de la quinta con el fin de evitar una excesiva carga que dañase la planta. Se contabilizaron los frutos raleados por árbol y se tuvieron en cuenta en el número final de frutos cosechados por árbol. A partir de 30 días post-caída de pétalos, en períodos de 20 días se midió el diámetro ecuatorial de 30 frutos al azar por planta hasta el momento de cosecha. En cosecha se midió rendimiento contabilizando el número de frutos y los kilos de fruta por árbol. A una muestra de 50 frutos al azar por árbol, se le midió el diámetro ecuatorial y se contabilizó el número de semillas totales y abortadas por fruto.
Análisis estadístico
Las variables ‘número medio de tubos polínicos’ y ‘distancia máxima recorrida por los tubos polínicos’, se analizaron a través de un Modelo Lineal Mixto, asumiendo distribución normal, y la diferencia de medias se analizó mediante el test de Tukey. Para comparar los efectos de los tratamientos sobre la germinación de polen se utilizó un Modelo Lineal Generalizado (McCullagh y Nelder, 1989), asumiéndose distribución binomial de la variable ‘número de granos de polen germinados en el total de granos evaluados’. En la comparación de medias se hicieron contrastes utilizando una prueba Chi2.
En el experimento realizado en condiciones de campo, las variables ‘número medio de semillas por fruto’ (totales, verdaderas y abortadas) y ‘calibre medio por fruto’ se analizaron a través de un Modelo Lineal Mixto, considerando el bloque como factor aleatorio, asumiendo distribución normal y la diferencia de medias se analizó mediante el test de Tukey. Se utilizó el paquete estadístico SAS, versión 9.1.3. (SAS Institute, Cary, NC, 2005), para el análisis estadístico de todas las variables medidas en campo y en laboratorio.
Resultados y discusión
Germinación de polen in vitro
El porcentaje de germinación de polen in vitro de ‘Montenegrina’ fue del 40% (Cuadro 1), que puede considerarse adecuado para los cítricos, de acuerdo a reportes de distintas especies y variedades, determinándose valores de entre 0,6% a 86,3%, los cuales difirieron entre especies y variedades de la misma especie y entre los dos años analizados (Pardo et al., 2007). En mandarina ‘Común’, variedad que dio origen a ‘Montenegrina’, reportaron un 67,9% a 73,5% de germinación de polen in vitro, utilizando la misma técnica de germinación y el mismo medio de cultivo que en el presente trabajo. El valor obtenido de germinación in vitro en nuestro trabajo, fue superior al reportado por Rodrígues y Cunha Dornelles (1999) en el cual los granos de polen de mandarina ‘Montenegrina’ presentaban un 96,13% de viabilidad y entre 19,36% y 33,68% de germinación en medio de cultivo con sacarosa, ácido bórico y nitrato de calcio.
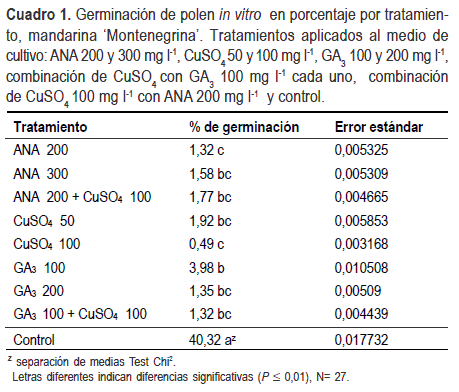
Todas las concentraciones y combinaciones de productos aplicados al medio de cultivo, inhibieron la germinación de polen in vitro en relación al control (Cuadro 1). En el caso del CuSO4, aplicado a 50 y 100 mg l-1, sólo o combinado con GA3 y ANA nuestros resultados en ‘Montenegrina’, confirman lo reportado por Mesejo et al. (2006) en mandarina ‘Fortune’, y se alinean con el efecto inhibidor de diferentes compuestos de cobre en la germinación del polen de diversas especies vegetales (Bristow y Shawa, 1981, Abott, 1991; Elezaby y Hasseeb, 1995). El cobre mineral ha demostrado ser polinicida en cerezo y damasco. A mayor concentración, menor porcentaje de germinación de polen y menor crecimiento de tubo polínico (Gür y Topdemir, 2008).
El GA3 sólo o combinado con CuSO4 alcanzó similares resultados (Cuadro 1). Se ha demostrado que las giberelinas actúan a nivel de los óvulos, ya sea mediante el aborto de los mismos, el desarrollo anormal del saco embrionario, o por el desarrollo desincronizado entre el estilo y el saco embrionario (Motomura e Ito, 1972, Fellman, et al.,1991). Nuestros resultados demuestran que el GA3 también afecta a los granos de polen inhibiendo su germinación en ‘Montenegrina’. Este resultado coincide con lo encontrado por Kimura et al. (1996) en la variedad de vid ‘Muscat Bailey’, donde la aplicación de GA3 solo o combinado con estreptomicina presentó los menores porcentajes de germinación de polen, mientras que la aplicación de estreptomicina sola no modificó el comportamiento de los testigos.
Las giberelinas han demostrado ser polinicidas en otras especies como Allium cepa, evitando el desarrollo de granos de polen y de anteras, produciendo plantas estériles (Van Der Meer y Van Bennekom, 1973, 1976).
El ANA redujo significativamente la germinación de polen in vitro confirmando evidencias en otras especies. Zhang y O’ Neill (1993) han propuesto que las auxinas, conjuntamente con el etileno actúan coordinando el desarrollo del ovario y del gametofito masculino para favorecer la fecundación. Se ha reportado que concentraciones mayores a 50 mg l-1 de auxina (AIA), inhibieron la germinación de polen en Allium cepa y provocaron detención del desarrollo del tubo polínico, el cual apareció como una mínima protuberancia (Kwan et al., 1969). Similares resultados han sido encontrados en manzano con la aplicación de AIA, verificándose germinación anormal de polen y efecto negativo en la elongación del tubo polínico (Elezaby y Hasseeb, 1995).
Germinación de polen y crecimiento de tubo polínico in vivo
En el día de aplicación de los tratamientos con las flores en estado de desarrollo 59 de la escala BBCH (Agustí et al., 1995), se observó muy poco polen a nivel del estigma. En el control germinaron en promedio sólo 18 granos de polen y ninguno de ellos creció más allá del estigma (Cuadro 2). Esto último indica que en estado de flor cerrada no existió posibilidad de autopolinización previa a la antesis.
A los cinco días post-aplicación, sólo el CuSO4 + GA3 y el ANA redujeron significativamente la germinación de polen, mientras que todos los tratamientos inhibieron el crecimiento del tubo polínico en relación al control (Cuadro 2). A los diez días post-aplicación en todos los tratamientos se verificó una disminución del número de tubos polínicos a nivel del estigma. En cuanto al crecimiento del tubo polínico, el ANA no provocó inhibición con respecto al testigo. Por el contrario, el GA3 fue el que presentó el mayor efecto inhibidor, deteniendo el crecimiento del tubo polínico en la zona del estigma, mientras que en esa fecha en el control, los tubos polínicos llegaron a recorrer más del 75% del estilo (Cuadro 2, Figura 1). Es importante señalar que estos resultados son nuevos para una especie autocompatible como ‘Montenegrina’, ya que los únicos reportes existentes sobre la inhibición del crecimiento del tubo polínico con GA3 en cítricos, son de Mesejo et al. (2008), en ‘Clemenules’, variedad autoincompatible.
El CuSO4, a los cinco y a los diez días post-aplicación redujo significativamente el crecimiento del tubo polínico. Esto coincide con lo observado en mandarina ‘Fortune’ donde la aplicación in vitro de 25 mg l-1 de CuSO4, ocho horas después de la germinación de polen detuvo por completo el crecimiento de tubo polínico, mientras que el testigo continuó creciendo y se incrementó un 50% en longitud (Mesejo et al., 2006). En ese mismo trabajo la aplicación de CuSO4 a flores en el momento de la polinización, también inhibió el crecimiento de tubo polínico lo cual coincide con los resultados obtenidos para mandarina ‘Montenegrina’. El cobre como mineral o bajo distintas formas químicas ha demostrado ser inhibidor del crecimiento del tubo polínico en varias especies de plantas (Abott, 1991, Sawidis y Reiss, 1995, Gür y Topdemir, 2008). El efecto del cobre mineral sobre el tubo polínico ha sido reportado, al igual que para otros metales pesados, afectando la estructura de la pared celular del tubo polínico (Speranza et al., 2007). En Lilium longiflorum se observó que el cobre mineral provocó una desorganización de la estructura de la pared celular a nivel del extremo del tubo, provocando así que no continuara su crecimiento (Sawidis y Reiss, 1995).
El GA3 aplicado previo a la antesis tuvo un efecto similar, inhibiendo el crecimiento del tubo polínico a los cinco y diez días post-aplicación. En ese período se interrumpió el crecimiento a nivel del estigma, provocando que no existiera interacción entre polen y estilo. Este resultado coincide con lo reportado por Mesejo et al. (2008), donde la aplicación de 10 mg l-1 de GA3 redujo el crecimiento del tubo polínico y sólo el 20% del estilo fue recorrido en flores de ‘Clemenules’ tratadas, contra un 60% en el control. El efecto dependió del momento de aplicación, ya que en las flores tratadas en antesis, los tubos polínicos no llegaron a los óvulos, y se produjo un arresto completo del crecimiento que impidió completamente la fertilización, a tal punto que la reducción en el número de semillas fue del 100% (Mesejo et al., 2008). Por otro lado en las flores tratadas el día posterior a la antesis, los mismos autores reportan que los tubos llegaron a fecundar el 15% de los óvulos doce días después y esto resultó en un 55% de reducción en el número de semillas. En el presente trabajo las aplicaciones fueron realizadas antes de la antesis. La inhibición del crecimiento del tubo polínico fue significativa en ambas fechas y se intensificó su efecto con el tiempo y la maduración del polen. Los mismos autores proponen que el GA3, además de tener un efecto directo inhibiendo la elongación del tubo polínico, también promueve la elongación del estilo haciendo que se dificulte aún más la llegada al óvulo. Esto podría ser una explicación al menor recorrido logrado por los tubos polínicos a los diez días, comparado con la distancia recorrida a los cinco días. Kimura et al. (1996) reportan resultados similares en la variedad de vid ‘Muscat Bailey’ con aplicaciones previas a plena floración, donde se pudieron observar muy pocos tubos polínicos y todos ellos sin sobrepasar la región estigmática. En una línea coincidente, en el cultivar de vid ‘Delaware’, aplicaciones prefloración de GA3, inhibieron significativamente el crecimiento de los tubos polínicos, impidiendo la penetración al micrópilo aún luego de tres y cinco días posteriores a la polinización. Se observó además que las aplicaciones en prefloración produjeron deformación del extremo del tubo polínico afectando su elongación y direccionamiento hacia el óvulo (Okamoto y Miura, 2005).
El ANA redujo significativamente el número de tubos polínicos en el estilo con respecto al control a los cinco y diez días post-aplicación. El crecimientode los mismos, también disminuyó a los cinco días post-aplicación y a la mitad a los diez días post-aplicación con respecto al testigo. Si bien las auxinas han demostrado ser reguladorasdel crecimiento de tubo polínico y del desarrollo de óvulo (Zhang y O’Neill, 1993), y concentraciones menores a 0,5 mg l-1 han promovido el crecimiento del tubo polínico en cebolla, las superiores a 50 y 100 mg l-1 han provocado su inhibición. Con 200 mg l-1 ni siquiera ocurrió la emergencia del tubo polínico, observándose apenas una pequeña protuberancia en el grano del polen (Kwan et al., 1969). Estos mismos autores han reportado que dosis superiores a 5 mg l-1 de AIA produjeron acumulación de dicho metabolito, lo cual provocó aumento del potencial osmótico que terminó con la ruptura del tubo polínico. En manzano, Elezaby y Hasseeb (1995) también reportan reducción del crecimiento de tubo polínico en plantas tratadas en plena floración con 10 y 100 mg l-1 de AIA con respecto a plantas sin tratar. Esa reducción del crecimiento del tubo polínico fue acompañada de una germinación anormal del grano de polen.
Inducción de partenocarpia en condiciones de campo
La aplicación simple y doble de GA3 logró disminuir significativamente el número de semillas verdaderas y totales en relación al control, con una reducción promedio de dos semillas por fruto (Cuadro 3). Este resultado es similar a lo reportado por Feinstein et al. (1975), donde una aplicación a principio y mitad de floración de 20 mg l-1 GA3 a árboles completos redujo el número de semillas desarrolladas por fruto en forma significativa de 12,5 a 9,7 y 10,3 promedio por fruto en mandarina ‘Dancy’, variedad autocompatible. Erner et al. (2008) reportaron resultados coincidentes con los anteriores, con dos aplicaciones de GA3 de 200 y 400 mg l-1 separadas por un período de dos semanas en pummelo ‘Chandler’ autoincompatible. La causa de no lograr una reducción más importante en el número de semillas, no parece ser la concentración aplicada, ya que el GA3 a 100 mg l-1 demostró ser el mejor tratamiento inhibidor de la germinación y el crecimiento del tubo polínico. La causa más probable está relacionada con los momentos de aplicación. En este trabajo se hicieron una o dos aplicaciones con un intervalo de quince días entre sí, la primera con 30-40% de flor abierta y la segunda con 70-80% de flor abierta. Con estas dos aplicaciones no se pudo cubrir un porcentaje importante de flores en los estados de mayor sensibilidad y por lo tanto muchas de ellas no fueron afectadas por los tratamientos. El momento de aplicación, relacionado al estado de desarrollo de las flores, es uno de los factores más importantes en la respuesta a la aplicación de GA3 tal como lo demostró Mesejo et al. (2008) en ‘Clemenules’. Las aplicaciones más tempranas, siete días preantesis, presentaron reducción del número de semillas del 52% y dos días preantesis la reducción fue del 26%. Pero la aplicación en antesis demostró ser la más efectiva, ya que produjo detención del crecimiento del tubo polínico, previniendo la fertilización y logrando una reducción del 100% en el número de semillas. ‘Montenegrina’ es una variedad autocompatible cuyas anteras abren al inicio de antesis, por lo que para que una aplicación sea eficiente en inducir partenocarpia, es necesario realizarla previo a la antesis, para evitar que exista germinación de polen a nivel del estigma. El número de semillas abortadas no presentó diferencias significativas entre tratamientos ni con el control (Cuadro 3). Este resultado coincide con los de Feinstein et al. (1975) donde aplicaciones con distintas dosis de GA3 tampoco produjeron diferencias significativas en el número de semillas abortadas entre sí, ni con el control.
Las aplicaciones simples y dobles de CuSO4 no provocaron reducción significativa del número de semillas verdaderas y totales por fruto comparado con el control (Cuadro 3). Este resultado no confirma el reporte de Mesejo et al. (2006), quienes verifican en dos años de trabajo una disminución significativa del número de semillas con aplicaciones de 25 mg l-1 de CuSO4 (aún menores a las aplicadas en el presente trabajo) en mandarina ‘Afourer’ en condiciones de polinización abierta, reduciendo el número de semillas por fruto en un 55% y 81% respectivamente. Erner et al. (2008) tampoco logran repetir esos resultados en pummelo ‘Chandler’ con aplicaciones de 25 y 100 mg l-1 de CuSO4. La causa podría ser que tanto ‘Chandler’ como ‘Montenegrina’, son variedades con baja capacidad partenocárpica y alta fertilidad de polen (Rodrígues y Cunha Dornelles, 1999, Yamamoto et al., 2006), lo cual hace que no respondan a la aplicación de CuSO4 de la misma forma que otras variedades que presentan alta capacidad partenocárpica.
Conclusiones
De todos los tratamientos inductores de partenocarpia el GA3 y el CuSO4 fueron los tratamientos más efectivos en inhibir la germinación de polen in vitro y el crecimiento del tubo polínico in vivo. El GA3 impidió el crecimiento del tubo polínico más allá de la zona del estigma a los diez días posteriores a la aplicación. El GA3 se presenta como el producto más efectivo para disminuir a niveles comercialmente aceptables el número de semillas por fruto en la variedad autocompatible ‘Montenegrina’.
Agradecimientos
Al Ing. Agr. MSc. Alegre Sasson y al Sr. Roberto Marrero por brindarnos las plantas, la maquinaria y la mano de obra necesaria para realizar las aplicaciones de campo. Al Laboratorio de Genética por permitir el uso del microscopio óptico con fluorescencia y al Laboratorio de Bioquímica de Facultad de Agronomía por brindar los materiales y su conocimiento en la preparación del medio de cultivo y del fluorocromo. Al INIA, a través de la beca INIA-UdelaR que brindó financiamiento para el presente trabajo.
Bibliografía
Abbott J.D. 1991. Fungicidal Inhibition of Pollen Germination and Germ-tube Elongation in Muskmelon. HortScience; 26(5): 529-530.
Agustí M., Zaragoza S., Bleiholder H., Buhr L., Hack H. y Klose R. 1995. Escala BBCH para la descripción de los estadios fenológicos del desarrollo de los agrios (Gén. Citrus). Lev. Agrícola; 332: 189-199.
Anderson C. 1996. Manual para productores de naranja y mandarina de la Región del Río Uruguay. INTA (Manual Serie «A» 2). 67 p.
Barry G. H. 2004. The Quest for Seedless Citrus Fruit. Proc. Int. Soc. Citriculture, 10 : 346.
Brewbaker J.L. and Kwack B. H. 1963. The Essential Role of Calcium Ion in the Pollen Tube Growth. Amer. J. Bot.; 50: 859-865.
Bristow P.R. and Shawa A. Y. 1981. The Influence of Fungicides on Pollen Germination and Yield of Cranberry.; J. Amer. Soc. Hort. Sci. 106(3): 290-292.
DIEA .2008. Encuesta Citrícola «Primavera 2007». Montevideo: MGAP (Serie de Encuestas Nº 259) 24p.
Domíngues M. C. e Rodrigues J. D. 2007. Redução de sementes do tangor ‘Murcott’ com a aplicação de biorreguladores durante o florescimento. Cienc. Agrotec., Lavras; 31 (3) :758-764.
Elezaby A. A. and Hasseeb G. M. 1995. Fungicidal Inhibition and Growth Regulator Promotion of Pollen Germination and Germ-tube Elongation in Apple. Acta Hortic.; 409: 179-183.
Erner Y., Tagari E. and Shmueli S. 2008. Reduction of Seed Number in Pummelo Chandler. Abstract XI Int. Citrus Congress, Wuhan, China. p. 82.
Feinstein B., Monselise S. P. and Goren R. 1975. Studies on the Reduction of Seed Number in Mandarins. HortScience; 10(4): 385-386.
Fellman C., Hoover E., Ascher P. D. and Luby J. 1991. Gibberellic Acid-induced Seedlessness in Field- Grown Vines of ‘Swenson Red’grapes. HortScience; 26 (7): 873- 875.
Gambetta G., Espino M., Pardo E., Alberti V., Arbiza H. and Gravina A. 2008. ‘Montenegrina’ Mandarin: Characterization of the Agronomic Behaviour and Fruit Size Improvement. Abstract Int. Citrus Congress, Wuhan, China. p. 84.
Gür N. and Topdemir A. 2008. Effects of Some Heavy Metal son in vitro Pollen Germination and Tube Growth of Apricot(Armenica vulgaris Lam.) And Cherry(Cerasus avium L.). World Applied Sciences Journal; 4(2): 195-198.
Kearns C. A. and Inouye D. W. 1993. Techniques for Pollination Biologists. Colorado: University Press of Colorado. 583p.
Kimura P., Okamoto G. and Hirano K. 1996. Effect of Gibberellic Acid and Streptomycin on Pollen Germination and Ovule and Seed Development in Muscat Bailey A. Am. J. Enol. Vitic.; 47 (2): 152 – 156.
Koehler-Santos P., Cunha Dornelles A. L. and Brandao de Freitas L. 2003. Characterization of Mandarin Citrus Germplasm from Southern Brazil by Morphological and Molecular Analyses. Pesq. Agropec. Bras., Brasilia; 38 (7): 797-806.
Kwan S. C., Hamson A. R., and Campbell W. F. 1969. The Effects of Different Chemicals on Pollen Germination and Tube Growth in Allium cepa. Amer. Soc. Hort. Sci.; 94: 561-562.
Lewin I. J. and Monselise S. P. 1976. Further Studies on the Reduction of Seeds in Mandarins by NAA sprays. Sci. Hortic.; 4: 229-284.
McCullagh P. and Nelder J. 1989. Generalized Linear Models. London: Chapman and Hall. 511p.
Mesejo C., Martínez-Fuentes A., Reig C. and Agustí M. 2008. Gibberellic Acid Impairs Fertilization in Clementine Mandarin under Cross-pollination Conditions. Plant Science; 175(3): 267-271.
Mesejo C., Martinez-Fuentes A., Reig C., Rivas F. and Agustí M. 2006. The Inhibitory Effect of CuSO4 on Citrus Pollen Germination and Pollen Tube Growth and its Application for the Production of Seedless Fruit. Plant Science; 170 (1): 37-43.
Motomura Y. and Ito H. 1972. Exogenous Gibberellin as Responsible for the seedless Berry Development of Grapes: II. Role and Effects of the Prebloom Gibberellin Application as Concerned with the Flowering, Seedlessness and Seedless Berry Development of Delaware and Campbell Early Grapes. Tohoku Journal of Agricultural Research; 23(1): 15-32.
Okamoto G. and Miura K. 2005. Effect of Pre-bloom GA Application on Pollen Tube Growth in cv. Delaware Grape Pistils. Vitis; 44(4): 157-159.
Pardo J., Bermejo A., Cano A. y Zaragoza S. 2007. La Germinación del polen y la formación de las semillas en los cítricos. Levante Agrícola; 384: 16-20.
Rodrígues L. e Cunha Dornelles A. 1999. Origem e caracterização horticultural da tangerineira ‘Montenegrina’. Laranja; 20 (1): 167-185.
Rodrígues L. R., Cunha Dornelles A. L. e Schifino-Wittmann M. T. 1999. Poliembrionia e número de sementes por fruto de quatro cultivares de tangerineira. Ciencia Rural, Santa María; 29(3): 469-474.
Sawidis T. and Reiss H. D. 1995. Effects of Heavy Metals on Pollen Tube Growth and Ultrastucture. Protoplasma; 185: 113-122.
Speranza A., Ferri P. Battistelli M., Falcieri E., Crinelli R. and Scoccianti V. 2007. Both Trivalent and Hexavalent Chromium Strongly Alter in vitro Germination and Ultrastructure of Kiwifruit Pollen. Chemosphere; 66: 1165–1174.
Stanley R.G. and Linskens H.F. 1974 Pollen: Biology, Chemistry, Management. New York: Springer. 307p.
Tominaga S. 1997. GA Sprays Delay and Reduce Physiological Fruit Drop in Ponkan Mandarin (Citrus reticulata Blanco). Acta Hortic.; 463: 301- 310.
Van der Meer Q. P. and Van Bennekom J. L. 1973. Gibberellic Acid as a Gametocide for the Common Onion (Allium cepa L.). Euphytica; 22: 239-243.
Van der Meer Q. P. and Van Bennekom J. L. 1976. Gibberellic Acid as a Gametocide for the Common Onion (Allium cepa L.) : II. The Effect of GA 4/7. Euphytica; 25: 293-296.
Yamamoto M., Kubo T. and Tominaga S. 2006. Self- and Cross-incompatibility of Various Citrus Accessions. J. Japan. Soc. Hort. Sci.; 75(5): 372-378.
Zhang X. S. and O’Neill S. D. 1993. Ovary and Gametophyte Development are Coordinately Regulated by Auxin and Ethylene Following Pollination. The Plant Cell; 5: 403-418.

