











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo

 Permalink
Permalink
1. Introducción
Los Hidrocarburos Aromáticos Policíclicos (HAP´s) constituyen un amplio grupo de compuestos químicos formados a partir de la combustión incompleta de materia orgánica. Se componen de dos o más anillos aromáticos que contienen cinco o seis átomos de carbono, siendo compuestos generalmente lipófilos e inestables fotoquímicamente. Existen más de 100 clases diferentes de HAP´s y se encuentran generalmente como mezclas complejas.1,2
Los HAP´s pueden entrar al organismo por diferentes vías: inhalatoria (humo de cigarrillo, de madera, de carbón y proveniente de aéreas industriales, gases de combustión en vehículos), digestiva (agua o alimentos), dérmica (contacto con partículas del suelo con altos niveles de HAP’s, o con aceite de motor). (3,4 El benzo(a)pireno (Bp) es el HAP más estudiado siendo clasificado por la Agencia internacional de investigación para el cáncer (IARC) como un carcinógeno del grupo I. (5
Sin tomar en cuenta a los trabajadores con exposición ocupacional, el consumo de cigarrillos y la ingestión de alimentos contaminados son la principal vía de exposición y una vez dentro del organismo se distribuyen en diversos órganos y tejidos que contienen grasa (ej.: riñones y tejido adiposo) y especialmente en el hígado. (1,4)
Un aspecto de interés para comprender la actividad biológica de los HAP´s es su metabolismo. (3 En organismos eucariotas los compuestos lipofílicos, tales como muchos productos naturales y compuestos sintéticos incluyendo drogas, carcinógenos y pesticidas, no pueden ser excretados a menos que se oxiden a compuestos más polares. (6
En humanos, el proceso de desintoxicación se lleva a cabo a través de un proceso de oxidación-conjugación que tiene como objetivo transformar estos compuestos para aumentar su comportamiento hidrosoluble, facilitando así la excreción urinaria. La producción de epóxidos, que son altamente reactivos, ocurre como un paso intermedio de la reacción de oxidación. Estos compuestos pueden unirse a moléculas complejas, como proteínas y ADN, generando así la genotoxicidad ligada a los HAP. (7
El proceso de detoxificación es realizado por enzimas del citocromo P450 las cuales están muy extendidas encontrándose en arqueas, bacterias y eucariotas. En Saccharomyces cerevisiae se ha descrito a la enzima 22-esterol desaturasa (CYP61, Erg5p) la cual además de la función de 22-desaturación durante la biosíntesis de ergosterol es también responsable de la metabolización y activación como promutágeno del Bp en ensayos de genotoxicidad. (6,8) Las lesiones producidas en el ADN por distintos agentes que perturban la estabilidad de esta macromolécula pueden repararse con cierta probabilidad, dependiendo de factores genéticos, metabólicos y ambientales. (9,10
Los aductos formados por la unión del derivado diol epóxido del Bp, inducen distorsión e inestabilidad en la cadena de ADN. La reparación por escisión de nucleótidos (NER) que se realiza con baja probabilidad de error es uno de los sistemas de defensa vital contra los aductos formados por los compuestos activos de los HAP´s que son potencialmente carcinógenos a nivel del ADN. En Saccharomyces cerevisiae la proteína Rad14 (Rad14p) (43 Kda) es crucial para la vía NER ya que la inactivación del gen RAD14 elimina por completo tanto la sub-vía que reconoce el daño en el genoma de forma global (GG-NER) como la sub-vía que reconoce el daño en aquellos genes que están transcribiéndose (TC-NER). (11,12
Además, los resultados de Chaurasia et al. (2013) demuestran el papel de un factor de reparación del ADN, Rad14p, en la estimulación de la transcripción además de su conocida función en NER, lo que implica a Rad14p como un importante regulador de la expresión génica. (13
Pero además del sistema NER se ha observado también una actividad importante de las vías de tolerancia al daño así como que en ciertas condiciones la exposición a Bp puede generar dobles roturas en el ADN. (14) Las vías de reparación que se activan frente a estas lesiones son la reparación por recombinación homóloga y la unión de extremos no homólogos. (15,16
Si éstos no pudieran repararse podría iniciarse la primera etapa de transformación maligna que podría llevar al desarrollo de cáncer. (10,17
Los polifenoles (PFs) constituyen uno de los grupos de metabolitos secundarios más numerosos y ubicuos de las plantas. (18) El ácido hidroxicinámico más abundante en los alimentos es el ácido clorogénico, el éster de ácido cafeico con ácido quínico (ácido 5-O-cafeoilquínico). (19
Estos compuestos están involucrados en los mecanismos de defensa de las plantas frente a agentes externos como la radiación ultravioleta y la agresión de patógenos Así mismo constituyen una parte importante de la dieta humana, por lo que también han despertado un gran interés desde el punto de vista nutricional. (20,21
La yerba mate (Ilex paraguariensis) presenta un alto contenido en PFs, que son reconocidos por su capacidad protectora a nivel celular y molecular frente a diferentes agentes genotóxicos. (22,23,24,25
En Uruguay una encuesta realizada por el Ministerio de Salud Pública (MSP) muestra que el 85% de la población mayor de 15 años consume por lo menos una vez por semana la infusión de yerba mate, siendo ésta una fuente para el consumo de ácido clorogénico entre otros antioxidantes. (26
Se ha demostrado que fenoles presentes en vegetales como los ácidos protocatéquico, clorogénico, tánico y el resveratrol, aplicados antes de HAP´s en piel de ratón reducen significativamente su unión al ADN epidérmico y los niveles de aductos de ADN. (27
El Bp produciría un aumento en los niveles de las enzimas CYP1A1/1A2 y CYP2B en animales tratados, así como de enzimas glutatión S-transferasa y NAD(P)H: quinona oxidorreductasa-1(NQO1), mientras que el pretratamiento con compuestos fenólicos antes de la aplicación de Bp disminuye significativamente las actividades de estas enzimas. (28
Por lo tanto, resulta de interés debido a la inevitable y continua exposición en nuestra vida diaria a estos compuestos y su reconocida capacidad cancerígena evaluar si el ácido clorogénico puede tener la capacidad de proteger a las células de la acción genotóxica que puede provocar el Bp utilizando como modelo eucariótico la levadura Saccharomyces cerevisiae.
La presente investigación propone que el ácido clorogénico podría proteger células de Saccharomyces cerevisiae salvajes y mutantes en el sistema de reparación por escisión de nucleótidos (NER) del daño inducido por benzo(a)pireno
2. Materiales y métodos
2.1 Cepas de levadura y condiciones de crecimiento.
Las siguientes cepas de Saccharomyces cerevisiae haploides fueron utilizadas en el presente análisis: SC7K lys2-3, cepa salvaje SX46A (MATa RAD ade2 his3-532 trp1-289 ura 3- 52) y su respectiva mutante SX46A rad14( (MATa rad14(∷HIS3 ade2 his3-532 trp1-289 ura 3-52). (29,30,31
La cepa SC7K lys2-3 se utilizó como biomarcador de mutagénesis además de para analizar la fracción de sobrevida y la cepa SX46A rad14( se utilizó por estar mutada en el gen que expresa la proteína Rad14 provocando la eliminación de la vía NER. De esta forma, poder evaluar la protección del ácido clorogénico ya que esta cepa no es capaz de reparar lesiones a través de la vía NER.
Las células de levadura se cultivaron hasta la fase exponencial (N = 1-2x107 células por mL) a 30 °C con aireación por agitación, en medio nutritivo líquido YPD: 1% extracto de levadura (Difco, US Biological), 2% peptona (Difco, US Biological) y glucosa al 2%(Sigma). (32
2.2 Agentes químicos.
Solución de ácido clorogénico (ACl). Se preparó una solución madre de ACl (Sigma) a una concentración de 0,9 mg/mL (PM: 354,31g/mol). Concentración final de 90 μg/mL (10v/v) en las diferentes alícuotas de cultivo celular (valor referencia en infusión de yerba mate). (33
Solución de Benzo(a)pireno (Bp). Se preparó una solución madre de Bp (Sigma) a una concentración de 2,5 μg/mL (PM=252,31 g/mol) y las diluciones se realizaron en agua destilada y esterilizada con el agregado de DMSO al 10 %.
Para las curvas dosis-efecto se realizaron tratamientos a diferentes concentraciones (10, 25, 50, 100, 200, 250 ng/mL), de acuerdo a observaciones previas. (34
2.3 Curva dosis efecto para Bp:
Las alícuotas de cultivo celular en medio de enriquecimiento (YPD) se incubaron por 60 minutos a 30°C con agitación a diferentes concentraciones de Bp (10, 25, 50, 100, 200, 250 ng/mL).
Se incubaron alícuotas de cultivo celular en medio de enriquecimiento durante 60 min a 30 °C con agitación adicionando Bp o ACl a una concentración final de 250 ng/mL y 90 μg/mL respectivamente.
Paralelamente, se realizaron tratamientos combinados de la misma forma que los individuales utilizando Bp y ACl a una concentración final de 250 ng/mL y 90 μg/mL respectivamente.
Inmediatamente después de los tratamientos, las suspensiones celulares se lavaron dos veces con agua destilada y se determinaron la fracción de supervivencia y la frecuencia mutagénica.
La fracción de supervivencia se determinó en función de la concentración de Bp con base en las curvas de supervivencia (Figura 1A ) y se seleccionó una concentración de 250 ng/mL para el tratamiento combinado. La elección de la dosis de Bp para los tratamientos simples y combinados se basa en los valores de fracción de sobrevida y frecuencia mutagénica obtenidos en las curvas respectivas, los cuales proporcionan un cambio sustantivo con respecto a los controles no tratados permitiendo observar cambios apreciables en el caso de que el ACl presentara protección.
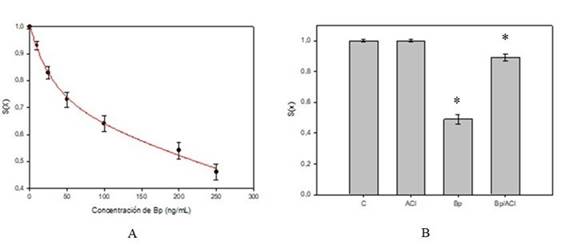
Figura 1: A, Fracción de sobrevida (S(x)) en función de concentraciones crecientes de Bp en la cepa SC7K lys2-3 con un ajuste exponencial decreciente doble, con 4 parámetros (y=a.e(-b.x)+c.e(-d.x)). B, Diagramas de barras que muestran la fracción de sobrevida (S(x)) en los siguientes tratamientos: control, simple (ACl o Bp) y combinado (Bp/ACl) para la cepa SC7K lys2-3. Datos de al menos tres experimentos reproducibles. (*p(0.05).
Alícuotas de cultivo celular se colocaron en placas con medio nutriente sólido. YPDA: YPD (1% extracto de levadura, 2% peptona y 2% de glucosa) + 2% agar (laboratorio Difco, USA) y se incubaron a 30°C durante 72 h. La fracción de sobrevida se calculó como: S(x,y) = Ns/No, donde Ns es el número de células sobrevivientes capaces de generar clones visibles/mL; No es el número total de células tratadas/mL; x es la concentración de Bp e y es la concentración de ACl.
2.4 Determinación de frecuencia mutagénica
Para determinar la frecuencia mutagénica, muestras celulares de SC7K lys2-3 se sembraron después del tratamiento en medios de omisión los cuales no contienen el aminoácido lysina (OM: 2% de dextrosa, 0,67% base nitrogenada de levadura (USBiological), 2% de agar) y se incubaron a 30°C por 21 días y se determinó el número de revertantes lys-LYS. La frecuencia mutagénica M (x, y) se calculó como: M(x, y) = Nm/Ns; donde Nm es el número de mutantes por mL, Ns es el número de células sobrevivientes por mL, x es la concentración de Bp, e y la concentración de ACl.
2.5 Determinación de las roturas de doble cadena en el ADN (DSBs)
Luego de ser expuestas a los tratamientos las muestras celulares se lavaron dos veces y se resuspendieron en buffer fosfato 0,5 M (pH 7,5) para una concentración de 5 x 108 células/mL. El ADN se aisló en bloques de agarosa de bajo punto de fusión tras el tratamiento enzimático (lyticasa, proteinasa K; Sigma, St Louis, MO). La separación cromosómica se llevó a cabo mediante electroforesis transversal de campo alterno (TAFE; Gene Línea II, Beckman, Fullerton, CA). Una vez terminado el tiempo de corrida el gel se colocó en el recipiente de tinción conteniendo bromuro de etidio (0.4 μg/mL) disuelto en buffer TAFE 1X durante 60 minutos a temperatura ambiente y protegido de la luz. Luego el gel se decoloró en buffer TAFE 1X conteniendo ribonucleasa A (1 μg/mL) (Sigma St Louis, MO) durante 3-4 h a 37°C protegido de la luz. 35,36 Posteriormente se colocó el gel en un transiluminador UV y se fotografió en forma digital. El análisis de las bandas de ADN se realizó mediante el software ImageJ (NIH, USA). En el caso de la inducción de rotura de doble cadena de ADN se observa una disminución en la intensidad de la banda correspondiente y un aumento concomitante en la fluorescencia intercalada entre bandas.
La estimación de los valores medios de roturas de doble cadena de ADN inducidas (DSB) y de las desviaciones estándar correspondientes se basaron en la distribución de Poisson como:
−ln Ix / Io,
donde Ix corresponde a la integral (superficie) de una determinada banda cromosómica en los densitogramas después del tratamiento e Io es la integral de la misma banda cromosómica en la respectiva muestra de control. (35,36,37,38
2.6 Análisis por Western blot.
Luego de realizar los tratamientos tanto simples como combinados, las muestras se centrifugaron para obtener un pellet de aproximadamente de 1 x 109 células.
La extracción de proteínas desnaturalizadas de levadura se realizó según el protocolo de Kushnirov, 2000. (39
Se realizó la electroforesis a 110 V fijo y amperaje libre durante 90 minutos y para confirmar tanto que la electroforesis se había realizado correctamente como que la concentración de proteínas en cada carril era igual se realizó la tinción con Azul de Comassie en uno de los geles. Para asegurar que todos los carriles tuvieran la misma cantidad de proteína, luego del control con azul de Coomassie, se procedió a tomar una captura de imagen en el escáner Odyssey y usar el programa ImageJ para medir las intensidades de las bandas encontradas a nivel de peso molecular de la proteína en estudio. El gel restante se utilizó para realizar la transferencia de proteínas.
El anticuerpo primario utilizado fue Anti-Rad14 (ab43577) de Abcam (a una dilución de 1/1000) y el anticuerpo secundario utilizado fue Goat anti- Mouse IRDye® 800 CW (926-32210) de LI-COR (a una dilución de 1/15000).
Las bandas de Rad 14p fueron visializadas usando Odyssey (LI-COR Biosciences) y se analizaron con Image Studio Lite (LI-COR Bioscience)
2.7 Análisis estadístico
La distribución binomial se aplicó al cálculo de los desvíos standard y errores standard en caso de los eventos binomiales: muerte - sobrevida y mutación - no mutación.
La distribución de Poisson se aplicó al cálculo y análisis del Nº de roturas dobles inducidas en el genoma. (25
Además se utilizó el test de t de comparación de dos medias independientes para estudiar si existían diferencias significativas entre los valores promedio de fracción de sobrevida y frecuencia mutagénica en las diferentes poblaciones estudiadas.
Y se utilizó ANOVA de una vía para la comparación de 3 medias independientes en el estudio de fracción de sobrevida y frecuencia mutagénica.
Para comparar las intensidades relativas de las bandas obtenidas en el Wester Blot se calculó el intervalo de confianza utilizando la distribución de T con 2 grados de libertad.
3. Resultados
3.1 Sobrevida celular
Para estudiar la relación de la fracción de supervivencia en función de dosis crecientes de Bp, se alicuotó el cultivo celular de SC7K lys2-3 y cada alícuota se incubó durante una hora con diferentes concentraciones del agente genotóxico. De esta relación dosis-efecto se seleccionó la concentración de 250 ng/mL para realizar los tratamientos combinados (Bp/ACl) así como los tratamientos simples de Bp y ACl. La alícuota de células que no recibió tratamiento sirvió como muestra de control.
La Figura 1A muestra la fracción sobreviviente en función de la concentración de Bp donde se puede observar que la solución a una concentración de 250 ng/mL induce una fracción de sobrevida de 0,46 ± 0,03 sobre la cepa SC7K lys2-3. También se puede observar que la relación entre la fracción de sobrevida y la dosis de Bp sigue un modelo exponencial decreciente doble con 4 parámetros (y=a.e(-b.x)+c.e(-d.x)) con un coeficiente de correlación R = 0,99.
La fracción superviviente de las células tratadas y sus correspondientes controles se muestran en la Figura 1 B. Se observó un aumento estadísticamente significativo en la sobrevida celular después del tratamiento combinado (Bp/ACl), en comparación con la sobrevida observada para Bp como agente único. Tanto las muestras control como las tratadas con ACl mostraron fracciones de sobrevida similares (1,0 ± 0,01).
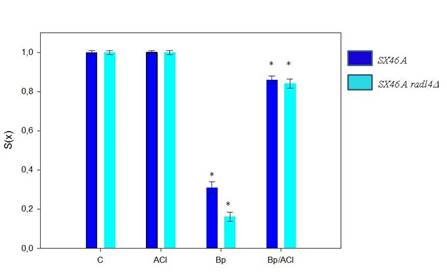
Al mismo tiempo, se realizó el análisis de sobrevida celular frente a los tratamientos simple y combinado en una cepa de levadura deficiente en el sistema de reparación por escisión de nucleótidos.
En las cepas SX46A y SX46A rad14( después de la exposición a Bp, las fracciones sobrevivientes fueron 0,31 ± 0,03 y 0,16 ± 0,02 respectivamente lo cual representa una diferencia significativa (Figura 2). Además, ambas cepas mostraron respuestas similares al tratamiento combinado de Bp y ACl observándose un aumento significativo de la sobrevida celular en estas condiciones con respecto al tratamiento con Bp como único agente.
3.2 Determinación de mutagénesis.
Para analizar el efecto diferencial de cada tratamiento sobre la mutagénesis inducida, muestras celulares de SC7K lys2-3 se sembraron en medio de omisión y se determinó la frecuencia de mutación en función de la concentración de Bp (Figura 3A). A partir de estos resultados se pudo observar un aumento exponencial en la frecuencia mutagénica en función de la concentración de Bp donde la relación entre las variables se ajustó a un modelo con 3 parámetros (y=y0+a.xb) con una correlación R=0,99 coeficiente (Figura 3A). Los análisis de frecuencia mutagénica de las muestras con tratamientos combinados mostraron una disminución significativa en la variable de estudio en comparación con las muestras tratadas solo con Bp (Figura 3B).
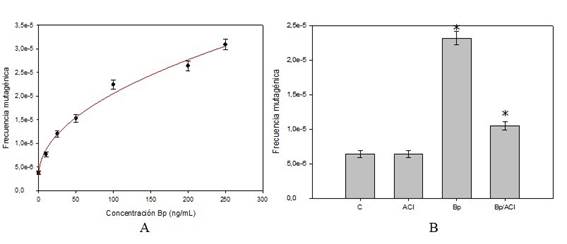
Figure 3: A, Frecuencia mutagénica en función de la concentración de Bp para la cepa SC7K lys2-3 con ajuste a un modelo de 3 parámetros (y=y0+a.xb). B, Muestra el marcado efecto antimutagénico del ACl en medio nutriente durante el tratamiento combinado Bp/ACl. Datos de al menos tres experimentos reproducibles. (*p(0.05).
Como puede observarse la M para el tratamiento con Bp como único agente fue de 2,32x10-5 ( 9,63x10-7 y para el tratamiento combinado Bp y ACl fue de 1,05x10-5 ( 6,63x10-7.
3.3 Determinación de DSBs.
Muestras celulares tratadas en las condiciones mencionadas anteriormente y los controles respectivos fueron sometidos a electroforesis de campos pulsados y alternos (TAFE) después del aislamiento de ADN.
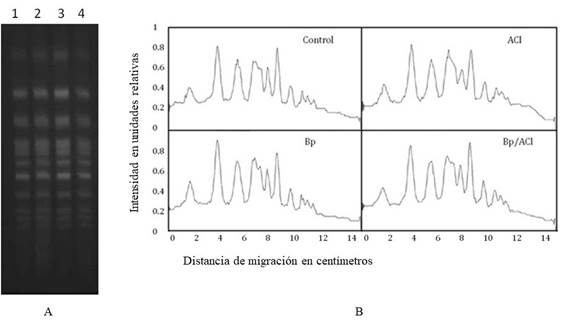
Se observó que en ninguna de las muestras analizadas se produjo fraccionamiento en el ADN.
Esto sugiere que, a la concentración utilizada en los experimentos (250 ng/mL), los daños que se producen en el genoma no implican la aparición de dobles roturas en la hebra de ADN. Esto puede observarse en la corrida electroforética (Figura 4A) y al comparar los perfiles densitográficos obtenidos para cada muestra (Figura 4B).
3.4 Expresión de la proteína Rad14.
Al realizar el análisis de Western Blot (WB) con un anticuerpo específico de Rad14 se observaron bandas muy tenues del peso molecular de la proteína en estudio en las condiciones control (Figura 5A carril 1 y 2).
En el carril correspondiente al tratamiento con Bp (Figura 5A carril 3) se observa una banda de mayor intensidad también en el peso molecular para la proteína en estudio. Éste aumento en la intensidad de la banda indica un aumento en el nivel de la proteína en estas condiciones.
En el caso del tratamiento combinado de Bp y ACl (Figura 5A carril 4) puede observarse una disminución en la intensidad de la banda lo cual sugiere que en estas condiciones la expresión de la proteína en estudio disminuye al compararla con el tratamiento con Bp como único agente. Acompañando al estudio cualitativo se realizó también una cuantificación de la diferencia de intensidades relativas de las bandas en cada muestra y se graficaron estos datos (Figura 5B).
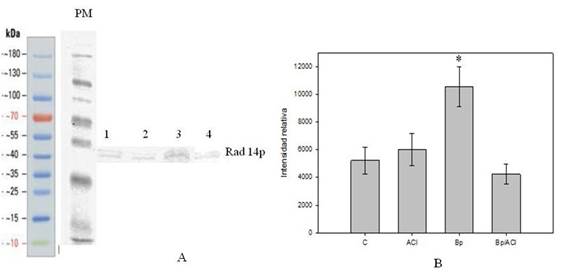
Figure 5: A, Western Blot (WB) (Anti-Rad14p) con las muestras de la cepa SC7K lys2-3 usando los siguientes tratamientos: MW, peso molecular; 1, Control; 2, ACl; 3, Bp; 4, Bp/ACl. Se denominó Rad14 la fila que representa el peso molecular correspondiente a la proteína en estudio. B, Intensidades relativas de las bandas correspondientes a Rad 14p para cada muestra. (*p(0.05)
4. Discusión
A partir de los resultados obtenidos se observa que el ácido clorogénico disminuye la letalidad y la actividad mutagénica producida por Bp, lo cual no se encuentra relacionado con un incremento en la remoción de las lesiones por el sistema NER. En el presente trabajo se observó un aumento de la letalidad celular con respecto a la curva de sobrevida en función de concentraciones crecientes de Bp y la relación entre las variables en estudio fue de tipo exponencial decreciente doble, con 4 parámetros (y=a.e(-b.x)+c.e(-d.x)) con un coeficiente de ajuste igual a R=0,99 (Figura 1A).
En los análisis de sobrevida realizados en tratamientos simples y combinados se pudo observar una disminución significativa de la letalidad celular en los tratamientos concomitantes con Bp y ACl (Figura 1B).
Cuando el análisis de fracción de sobrevida se realizó en cultivos de una cepa deficiente en la proteína Rad14 se observó una disminución de la fracción de sobrevida en las muestras tratadas con Bp como único agente en comparación con la muestra control. Por otro lado, cuando el tratamiento fue en combinación con Bp y ACl se observó una disminución significativa de la letalidad (Figura 2).
Es importante resaltar que la fracción de sobrevida no se alteró significativamente en los tratamientos con ACl como único agente en comparación con la muestra control.
Cuando se realizaron estudios de frecuencia mutagénica en función de concentraciones crecientes de Bp se observó un aumento significativo ajustándose a un modelo con 3 parámetros (y=y0+a.xb) y presentando un coeficiente de ajuste de R= 0,99 (Figura 3A).
Se observó además que la frecuencia de mutación espontánea no se alteró significativamente en el tratamiento control con ACl como único agente a la concentración de trabajo.
Así mismo se observó una disminución significativa de la fracción mutagénica en los tratamientos combinados con Bp y ACl (Figura 3B).
Los aductos formados por los productos reactivos de los HAP´s son procesados en su mayoría a través de la vía NER, la cual como se mencionó anteriormente es una vía de reparación libre de error. Por lo tanto el nivel de frecuencia mutagénica que presentan las muestras tratadas con Bp puede deberse a que otras vías de reparación propensas a error están procesando parte de las lesiones producidas.
La generación de la frecuencia mutagénica observada puede deberse a que parte de las lesiones se procesan a través de la vía de síntesis a través de la lesión (TLS por sus siglas en ingles). (40 Cuando la célula ingresa en la fase S la presencia de aductos voluminosos en el ADN inhiben las polimerasas de alta fidelidad, bloquean las horquillas de replicación que conducirían a la muerte celular y la vía TLS (dependiente de RAD6) recluta al lugar donde la replicación se estancó polimerasas de baja fidelidad. (40
Sería importante estudiar a futuro como se modulan las proteínas vinculadas a estas vías de reparación frente a la exposición a Bp.
En cuanto al análisis del fraccionamiento cromosómico los datos muestran que a la concentración de Bp utilizada en este trabajo no se generan dobles roturas de ADN con lo cual no se pudo evaluar si el ACl presenta alguna acción protectora contra este tipo de daño (Figura 4 A y 4B ).
Esta afirmación surge del estudio de la relación entre las amplitudes de los picos así como la relación de la distancia recorrida en las abscisas para los diferentes picos. Al no existir diferencias significativas entre estas medidas se puede descartar la posibilidad de una adhesión entre los fragmentos de ADN (rearreglos) que no permitiera evidenciar la fragmentación (Figura 4 B).
La elección de utilizar 60 minutos de incubación se debe a dos motivos, uno de ellos es la elección de trabajar dentro de un tiempo menor al tiempo de generación celular de esta cepa de levaduras. Ademad en experimentos previos con tiempo de incubación de 40 minutos no se observaron diferencias ni en la fracción de sobrevida ni en la frecuencia mutagénica de las muestras tratadas con el compuesto Bp y la muestra control.
Tanto los datos obtenidos, que muestran una disminución en la letalidad celular y en la frecuencia mutagénica en presencia de ACl, así como las observaciones que muestran que no se altera la sobrevida ni la frecuencia mutagénica espontánea en presencia de ACl como único agente, están en sintonía con el comportamiento de otros polifenoles referenciados en la bibliografía consultada. (41,42,43,44,45
Los resultados de este trabajo indican que el ACl, a una concentración igual a la encontrada en la infusión de yerba mate, tuvo un efecto protector significativo contra el daño producido por Bp en poblaciones celulares haploides de S. cerevisiae.
En aquellas muestras que fueron tratadas con Bp como único agente en el análisis de Western Blot, se observó un aumento en la expresión de la proteína Rad14, uno de los pasos cruciales en la activación de la vía NER (Figura 5 A y 5B ).
En las muestras que fueron tratadas concomitantemente con Bp y ACl, se observó una disminución del nivel de la proteína Rad14 lo cual indicaría que las distorsiones provocadas en el ADN por el Bp han sido inhibidas.
El ácido clorogénico disminuyó de forma significativa tanto la letalidad como la frecuencia de mutagénesis en todos los tratamientos.
El hecho que la cepa deficiente en la proteína Rad14 no mostró diferencias significativas entre todos los tratamientos en presencia de ACl, indica que la modulación de los daños producidos por el ACl no estaría vinculada con la vía de reparación NER.
Como ya hemos mencionado los HAP´s provocan daño en el ADN al ser activados a través de las reacciones de metabolización por las enzimas de citocromo P450. Los productos diol-epóxidos que se producen en estas reacciones se podrían unir covalentemente a las bases del ADN produciendo una gran distorsión espacial de la doble hebra.
En conclusión, la protección del ácido clorogénico observada en la disminución de la letalidad y la actividad mutagénica producida por Bp no se vio relacionada con un incremento en la remoción de las lesiones por el sistema NER.
La forma en que el ACl provoca esta protección puede estar vinculada a diferentes acciones como son una acción atrapadora del compuesto reactivo del Bp, una acción inhibitoria sobre la expresión de las enzimas que metabolizan este tipo de compuesto o también alguna acción que module la expresión de los genes que codifican para las mismas enzimas. Estas presunciones pueden ser semillas de futuras investigaciones que intenten dilucidar a qué nivel o niveles actúa el ACl como agente protector.
El presente trabajo presenta algunas limitaciones, se debe tener en cuenta que algunos resultados son de tipo preliminar, considerandose relevante profundizar estos estudios sobre los mecanismos de interacción o de modulación que se da a través de la acción del ACl. Uno de los puntos débiles del artículo se relaciona con la metodología del análisis de proteínas. En nuestro caso no se utilizó un método bioquímico para medir las concentraciones de proteínas antes de realizar la electroforesis en el análisis por Western blot. Esto implica que aunque se hayan medido las intensidades de las bandas en el gel teñido con azul de Coomassie los resultados son de tipo cualitativo. Se proyecta profundizar en estos análisis de manera de confirmar los datos preliminares.

