










Serviços Personalizados
Journal

Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Links relacionados
Compartilhar

 Permalink
PermalinkOdontoestomatología
versão impressa ISSN 0797-0374versão On-line ISSN 1688-9339
Odontoestomatología vol.20 no.32 Montevideo dez. 2018
https://doi.org/10.22592/ode2018n32a10
Research
Cell proliferation study in human tooth germs
1
 http://orcid.org/0000-0001-7747-6718
http://orcid.org/0000-0001-7747-6718
2
http://orcid.org/0000-0002-0617-1279
3
http://orcid.org/0000-0003-1137-6866
1 Área de Patología Molecular Estomatológica, Facultad de Odontología, Universidad de la República, Uruguay. vanesapereira91@hotmail.com
2 Área de Patología Molecular Estomatológica, Facultad de Odontología, Universidad de la República, Uruguay.
3 Área de Patología Molecular Estomatológica, Facultad de Odontología, Universidad de la República, Uruguay
4 Área de Patología Molecular Estomatológica, Facultad de Odontología, Universidad de la República, Uruguay.
5 Cátedra de Histología, Facultad de Odontología, Universidad de la República, Uruguay.
The aim of this study was to determine the expression of MCM4-5-6 in human tooth germs in the bell stage.
Materials and methods:
Histological samples were collected from four fetal maxillae placed in paraffin at the block archive of the Histology Department of the School of Dentistry, UdelaR. Sections were made for HE routine technique and for immunohistochemistry technique for MCM4-5-6.
Results:
Different regions of the enamel organ showed 100% positivity in the intermediate layer, a variation from 100% to 0% in the inner epithelium from the cervical loop to the incisal area, and 0% in the stellar reticulum as well as the outer epithelium.
Conclusions:
The results show and confirm the proliferative action of the different areas of the enamel organ.
Keywords: MCM4; MCM5; MCM6; tooth germ; cell proliferation
El objetivo de este estudio fue conocer la expresión de MCM4-5-6 en gérmenes dentarios humanos en estado de campana.
Materiales y Métodos
Se obtuvieron preparados histológicos de 4 maxilares fetales incluidos en parafina en el archivo de bloques de la cátedra de Histología de la Facultad de Odontología, UdelaR. Se procedió al corte de los mismos en secciones para técnica de rutina (HE) y de IHQ para MCM 4, 5 y 6.
Resultados:
Las diferentes regiones del órgano del esmalte mostraron 100 % de positividad en el estrato intermedio, una variación de 100 % a 0 % en el epitelio interno del órgano del esmalte, desde el sector cervical al sector incisal del mismo, y0% tanto en el retículo estrellado como en el epitelio externo del órgano del esmalte.
Conclusiones:
Los resultados obtenidos permitieron evidenciar y confirmar la acción proliferativa de las diferentes zonas del órgano del esmalte.
Palabras clave: mcm4; mcm5; mcm6; gérmen dentario; proliferación celular
Introduction
Tooth organogenesis is a process involving a wide range of cellular functions which include, alternately, the participation of ectomesenchymal and epithelial elements, represented by the dental papilla and the enamel organ, respectively1.During tooth development, the germ undergoes different morphological stages, including the bud stage, the cap stage and the early and advanced bell stage1. Regarding structure, the tooth germ has three parts: two of ectomesenchymal origin (the papilla and the dental sac or follicle), and an epithelial one (the enamel organ)2. Dental papilla is responsible for the formation of pulp and dentin, both on the crown and the root. In turn, the dental sac or follicle is responsible for the development of the deep periodontium components and the connective elements of the superficial periodontium. Finally, the enamel organ participates in enamel formation, especially at the start of the inner epithelium3. All these physiological processes occur, among other things, through the cellular proliferation that results from an increase in the number of cells because of cell division4.
There are various cell proliferation markers, including the MCM2-7 complex (minichromosome maintenance)5.The MCM2-7 complex is formed by six different proteins (MCM2, MCM3, MCM4, MCM5, MCM6, MCM7) and is part of the pre-replication complex of the cell cycle, so it has an essential role in DNA replication6-7. One of its roles is to start the replication process at the right time, for which it must change its three-dimensional structure. When inactive, the MCM2-7 complex has a reversible discontinuity in its toroidal structure. As MCM2 and MCM5 bind, they close this discontinuity, which allows the MCM2-7 complex to activate8-9. It is also responsible for helicase activity10, stops replication11, verifies that all the DNA is replicated12, and prevents DNA from replicating more than once in the same cell cycle13. The proteins in this complex are expressed throughout the cell cycle from the early G1 phase onwards, as well as in cells with proliferative potential, but not in quiescent cells (G0)14.
This work aims to study the proliferative activity of human tooth germs in the bell stage through the expression of molecules from the MCM4-5-6 complex in the outer epithelium, stellate reticulum, intermediate stratum and inner epithelium of the enamel organ.
Materials and methods
We studied eight human tooth germs classified in the bell stage from the archive of the Histology Department, School of Dentistry, University of the Republic, Montevideo, Uruguay. They were obtained from four fetal maxillae placed in paraffin blocks of between 17 and 24 weeks of development. They are listed as processed in 1970 and were cut with a microtome, in 4-micron sections, and placed on silanized plates. A section was taken per sample to be stained with the hematoxylin and eosin technique, as well as three sections for immunohistochemistry (IHC) for MCM4-5-6 proteins.
For the immunohistochemical technique, the sections were deparaffinized in a stove at 45ºC for 60 min and subsequently left in xylol for 5 minutes. They were then hydrated with decreasing concentrations of absolute alcohols (90%, 80%, 70% and 50%) and rinsed twice with distilled water. To unmask epitopes, the antigen retrieval technique was used with a 10mM (pH 6.2) sodium citrate solution using a pressure cooker in microwave oven at full power (750 W) for 6 min. Then they were left to cool at room temperature and subsequently rinsed with distilled water. Endogenous peroxidase were blocked with hydrogen peroxide (0.9%) for 5 minutes, followed by washes with distilled water and pH 7.4 phosphate buffered saline (PBS). The slides were placed on plates and then in racks for immunohistochemistal tests by capillary action. We incubated the monoclonal primary antibodies against MCM4-5-6 (Leica Biosystems, Newcastle, UK, dilution 1:100) for 45 minutes. The sections were then incubated with the second biotinylated anti-mouse/anti-rabbit antibody and with the streptavidin/peroxidase complex (LSA-B + Labeled streptavidin-biotin, DakoCorporation, Carpinteria CA, USA) for 30 minutes each. The products of the reaction were visualized with substrate 3.3’-diaminobenzidin-H2O2 (Dako Corporation, Carpentry, CA, USA). The sections were counterstained with Mayer’s hematoxylin. Ameloblastoma specimens were used as blank samples.
The positive cells in the outer epithelium, stellate reticulum and intermediate stratum were counted, as well as in the cervical, middle and apical thirds of the inner epithelium of the enamel organ, according to the method described by Bologna et al.15.
We evaluated the expression of each antibody in selected areas rich in epithelial cells. Five microphotographs were taken per case, with 40x magnification in each field, with an Olympus® light microscope. The image was converted to JPEG format. Each file was opened in Microsoft PowerPoint, and with the “table” function, we created a 10x10 table which was superimposed on the microphotograph to facilitate the count. The positive and negative cells were counted manually. The rule of three was applied (total of positive cells x100/total number of cells). The percentage of positive cells was calculated for each field.
Results
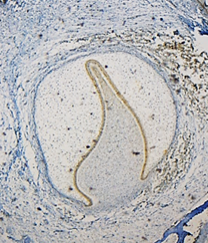
Once the specimens had been analyzed, we noted that the distribution of cell positivity in each of the thirds of the inner epithelium of the enamel organ was uniform for all the components of the MCM complex studied (Fig.1).
However, these cell proliferation markers are not expressed in the outer epithelium cells of the enamel organ and of the stellate reticulum, but express 100% positivity in the intermediate stratum cells.
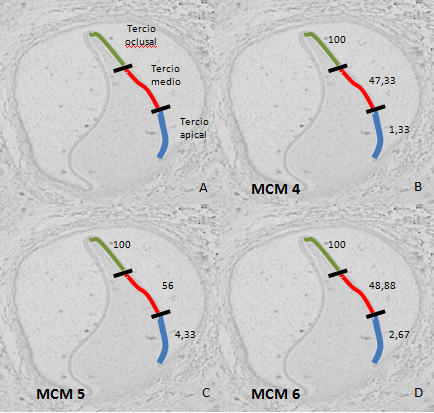
The peripheral section of the dental papilla presents a reverse distribution for the three markers, showing a continuum of lower proliferation rate in the cervical portion of the papilla, somewhat greater in the middle third and a high positivity rate in the third incisal or occlusal plane (Figs. 2 and 3).
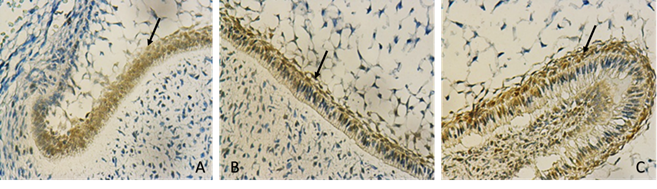
Discussion
Immunodetection of proteins involved in the process of cell proliferation is usually performed using biomarkers such as Ki-67, which is considered the gold standard16. This paper aims, for the first time, to identify three cell proliferation markers not studied so far in tooth germs: MCM4, 5 and 6. One of the differences between this complex and protein Ki-67, among others, is that protein Ki-67 is not expressed in the early G1 stage but from late G1, and continues throughout the cell cycle, except in quiescent cells (G0). The MCM complex molecules are expressed even in early G1, and are considered more sensitive in terms of their molecular detection17-18.
Previous studies conducted by Muica et al. in 2014 determined that Ki-67 expression decreases with the progress of the tooth development stages in all tissues and strata that make up the tooth germ19. Kero et al. support these results after their study conducted in the same year, where they determined that increased cell proliferation allows the enamel organ to grow, while its selective reduction in the different components of the germ leads to the disintegration of certain dental structures20. The fall of the cell proliferation index coincides with the beginning of the ameloblast and odontoblast differentiation20. In turn, each cell layer that makes up the tooth germ has specific proliferation activity. The proliferation rate of the inner epithelium of the enamel organ and the dental papilla peak in week 14, falling in week 21 during the late bell stage. This coincides with the appearance of mature enamel in the crown area, or ameloblast and odontoblast differentiation20. Additionally, both the outer epithelium and the stellar reticulum showed a steady decline in the proliferation activity throughout this stage. The distribution of proliferating cells was more uniform at the dental papilla in all the stages studied, compared to the rest of the cell layers20.
Comparing our results with the studies cited, in our study there is a gradual decrease in the MCM4, 5 and 6 percentage in the epithelium of the enamel organ, which might indicate a decline in the proliferation to promote selective tooth disintegration. This makes it possible to move to a more mature state of the tissue, which coincides with the differentiation of ameloblasts and odontoblasts. This also presents the negative expression of markers at the outer epithelium and the stellar reticulum.
Although MCM markers are more sensitive in certain tissues compared to the action of Ki-6717-18, we believe that these studies can be compared to our results.
Studies based on the invagination patterns of the Hertwig epithelial root sheath and the mesial root formation, as well as the formation of the tooth bifurcation, determine that these regions are correlated with the condensation of highly proliferative mesenchymal cells. In molars, the percentage of Ki-67 was 50% higher than in the mesial root areas with bifurcation, which suggests that pulp proliferation induces epithelial migration at this stage21-22.
Therefore, based on our results, where the positive expression of these three MCM proteins decreases (100% at the incisal level, 47-56% in the middle and reaching 1.33-4.33% at cervical level in the early bell stage), we believe that as odontogenesis stages progress jointly with cell differentiation, the different strata of tooth formation express levels of cell proliferation in a differential manner, with an epithelial-mesenchymal induction, concluding with the normal development of teeth.
Conclusions
According to the data obtained, we can consider that the proteins of the MCM Complex 4, 5 and 6 are indicators of proliferative activity in the process of ondontogenesis. Our findings show a continuum of concentrations of these molecules with marked differences between the cervical, middle and crown sections of the inner epithelium of the enamel organ. This is consistent with the cellular activity described by several authors in the literature. Such evidence is also found in the cells of the dental papilla, which should be the focus of further research
Referencias
1. Mass R, Bei M. The genetic control of early tooth development. Crit. Rev. Oral Biol. Med. 1997; 8 (1): 4-39. [ Links ]
2. Nanci A. Ten Cate's Oral Histology. 8th. ed. London: Elsevier, 2013. [ Links ]
3. Thesleff I, Keränen S, Jernvall J. Enamel knots as signaling centers linking tooth morphogenesis and odontoblast differentiation. Adv. Dent. Res. 2001; 15: 14-18. [ Links ]
4. Alver RC, Chadha GS, Blow JJ. The contribution of dormant origins to genome stability: from cell biology to human genetics. DNA Repair. 2014; 19 (100): 182-9. [ Links ]
5. Forsburg SL. Eukaryotic MCM proteins: beyond replication initiation. Microbiol Mol Biol Rev. 2004; 68 (1), 109-131. [ Links ]
6. Laskey R. The Croonian Lecture 2001 hunting the antisocial cancer cell: MCM proteins and their exploitation. Philos Trans R Soc Lond B Biol Sci. 2005; 360 (1458): 1119-1132. [ Links ]
7. Simon NE, Schwacha A. The Mcm2-7 replicative helicase: a promising chemotherapeutic target. BioMed Research International. 2014; 2014: 14. [ Links ]
8. Tanaka S, Umemori T, Hirai K, Muramatsu S, Kamimura Y, Araki H. CDK-dependent phosphorylation of Sld2 and Sld3 initiates DNA replication in budding yeast. Nature. 2007; 445 (7125): 328. [ Links ]
9. Zegerman P, Diffley JF. DNA replication as a target of the DNA damage checkpoint. DNA repair (Amst). 2009; 8 (9): 1077-1088. [ Links ]
10. Byun TS, Pacek M, Yee MC, Walter JC, Cimprich KA. Functional uncoupling of MCM helicase and DNA polymerase activities activates the ATR-dependent checkpoint. Genes Dev. 2005; 19 (9): 1040-1052. [ Links ]
11. Cortez D, Glick G, Elledge SJ. Minichromosome maintenance proteins are direct targets of the ATM and ATR checkpoint kinases. Proc Natl Acad Sci USA. 2004; 101 (27): 10078-10083. [ Links ]
12. Zhai Y, Li N, Jiang H, Huang X, Gao N, Tye BK. Unique Roles of the Non-identical MCM Subunits in DNA Replication Licensing. Mol Cell. 2017; 20;67(2): 168-179. [ Links ]
13. Deegan TD, Diffley JF. MCM: one ring to rule them all. Curr Opin Struct Biol. 2016; 37: 145-51. [ Links ]
14. Ishimi Y, Okayasu I, Kato C, Kwon HJ, Kimura H, Yamada K, Song SY. Enhanced expression of Mcm proteins in cancer cells derived from uterine cervix. Eur J Biochem. 2003; 270 (6): 1089-1101. [ Links ]
15. Bologna-Molina R, Damián-Matsumura P, Molina-Frechero N. An easy cell counting method for immunohistochemistry that does not use an image analysis program. Histopathology. 2011; 59 (4): 801-3. [ Links ]
16. Pontes HA, Pontes FS, Silva BS, Cury SE, Fonseca FP, Salim RA, Pintor Júnior Ddos S. Imunoexpression of ki67, proliferativecell nuclear antigen, and Bcl-2 proteins in a case of ameloblasticfibrosarcoma. Ann DiagnPathol. 2010; 14 (6): 447-52. [ Links ]
17. Scholzen T, Gerdes J. The Ki-67 protein: fromtheknown and theunknown. J Cell Physiol. 2000; 182: 311-22.33. [ Links ]
18. Kodani I, Osaki M, Shomori K, Araki K, Goto E, Ryoke K, Ito H. Minichromosome maintenance 2 expression is correlated with mode of invasion and prognosis in oral squamous cell carcinomas. J Oral PatholMed. 2003; 32 (8): 468-74. 34. [ Links ]
19. Muica Nagy-Bota MC, Pap Z, Denes L, Ghizdavat A, Brînzaniuc K, LupCosarca AS, ChibeleanCires-Marginean M, Pacurar M, Pávai Z. Immunohistochemical study of Ki67, CD34 and p53 expression in human tooth buds. Rom J MorpholEmbryol. 2014; 55 (1):43-8. [ Links ]
20. Kero D, Govorko D K, Vukojevic K, Cubela M, Soljic V, Saraga-Babic M. Expression of cytokeratin 8, vimentin, syndecan-1 and Ki-67 during human tooth development. Journal of molecular histology, 2014; 45 (6): 627-640. [ Links ]
21. Sohn WJ, Choi MA, Yamamoto H, Lee S, Lee Y, Jung JK, Jin MU, An CH, Jung HS, Suh JY, Shin HI, Kim JY. Contribution of Mesenchymal Proliferation in tooth root Morphogenesis. J Dent Res. 2014; 93 (1): 78-83. [ Links ]
22. Iwamoto T, Nakamura T, Ishikawa M, Yoshizaki K, Sugimoto A, Ida-Yonemochi H, Masahiro Saito, Yoshihiko Yamada, Satoshi Fukumoto. Pannexin 3 regulates proliferation and differentiation of odontoblasts via its hemichannel activities. https://doi.org/10.1371/journal.pone.0177557 [ Links ]
Received: May 14, 2018; Accepted: August 09, 2018
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
