










Servicios Personalizados
Revista

Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Links relacionados
Compartir

 Permalink
PermalinkOdontoestomatología
versión impresa ISSN 0797-0374versión On-line ISSN 1688-9339
Odontoestomatología vol.18 no.28 Montevideo nov. 2016
Metalloproteinases (MMPs) of the extracellular matrix in Dentistry
Pereira Prado, Vanesa*, Asquino, Natalia**, Apellaniz, Delmira***, Bueno Rossy, Luis****, Tapia, Gabriel*****, Bologna Molina, Ronell******
* Teaching Assistant, Molecular Pathology in Stomatology. Dental Sciences MSc student. School of Dentistry, Universidad de la República. Uruguay.
vaneapereira91@hotmail.com
** Teaching Assistant, Periodontics Department. MSc student (PROINBIO). School of Dentistry, Universidad de la República. Uruguay.
*** Dental Sciences MSc student. School of Dentistry, Universidad de la República. Uruguay.
**** Professor at the Periodontics Department. Director of Postgraduate Degree in Periodontics, School of Dentistry, Universidad de la República. Uruguay.
***** Associate Professor, Histology Department, School of Dentistry, Universidad de la República. Uruguay.
****** Professor at the Molecular Pathology in Stomatology Area, School of Dentistry, Universidad de la República. Uruguay.
Abstract
Most body tissues are found in a constant renovation process, modulated by a large number of proteins that induce tissue synthesis and degradation over time. Matrix metalloproteinases (MMPs) are a family of 25 zinc-dependent endopeptidases in charge of the cleavage of the immediate components of the extracellular matrix. Certain MMPs, especially MMP-2 and MMP-9, are related to the development of dental germ as well as odontogenic tumor invasion and proliferation. They are a significant factor in tumor aggressiveness and possible treatments. Moreover, both MMPs participate in the progression of dental caries and apical periodontitis in patients with necrotic pulp. Several periodontal pathogens promote the secretion of both MMP-2 and MMP-9, increasing the destruction of periodontal tissues, which is closely linked to the progression of the disease. High levels of these MMPs are found in patients undergoing orthodontic treatment, not only in the compression side but also on the tension one. This establishes a connection between tooth movement and tissue renovation. This review analyzes MMPs expression and usefulness in different areas of dental science, emphasizing their potential as biomarkers.
Keywords: Matrix metalloproteinases, Extracellular matrix, Dentistry.
Received on: 10 Dec 15 – Accepted on: 11 Aug 16
Introduction
Several types of cells that produce a wide variety of macromolecules, among them, various proteins, are involved in the development of dental germs. Some of these proteins worth noting are matrix metalloproteinases (MMPs), which play a role in the degradation of the extracellular matrix and whose functions are coordinated with tissue inhibitors of metalloproteinases called TIMPs. Certain MMPs have been linked to the progression of tumor pathologies, metastases and tumor aggressiveness, while some have been used as markers to help in the diagnosis and prognosis in various tumors of epithelial origin. MMP-2 can degrade enamel proteins such as amelogenin and, consequently, promote a dental tissue that is more prone to cariogenic attacks (1). Apical lesions, caused after pulp necrosis, mark an increase in the activity of MMP-2 and MMP-9, which could contribute to the progression of apical periodontitis (2). Since studies have shown an increased expression and activity of MMP-9 in patients with periodontitis, it could be considered a predictive indicator of the disease (3). Several MMPs, among them MMP-2 and MMP-9, have been proven to be in higher levels in compression and tension sites of teeth subjected to orthodontic movements (4). The aim of this paper is to gather information on the expression of said proteins in odontogenic tumors and in tooth germs, as well as their role in restorative dentistry, endodontics and periodontics.
Methodology used for collecting data
A computer search of the literature in Spanish and English was carried out using the Pubmed, SciELO, Science Direct and Timbó (portal created by the National Research and Innovation Agency of Uruguay that provides access to literature) databases for the 2005-2015 period, using keywords such as: “metaloproteinasas, matriz extracelular, odontología, matrix-metalloproteinases, extracellular-matrix, dentistry”, and 29 papers from the last 6 years were selected to guide this literature review.
What are matrix metalloproteinases?
MMPs are integral proteins that are part of the extracellular matrix (ECM) and can activate growth factors, within their immediate environment, cell surface receptors and adhesion molecules (5). These metalloproteinases constitute an important family of zinc-dependent endopeptidases and their activity is regulated by specific inhibitors known as tissue inhibitors of metalloproteinases (TIMPs) (5).
Twenty-five members of the MMP family have been described and classified into five subfamilies according to their function and structure: collagenases, gelatinases, stromelysins, membrane-type matrix metalloproteinases (MT-MMPs) and other MMPs (5).
The structure of MMPs comprises several domains shared by most of them, which are necessary for the intracellular movement of the enzyme to the membrane, and which are eliminated after the protease is secreted, as well as a domain with latent enzymatic capacity (6).
Functions of the matrix metalloproteinases
MMPs are a group of enzymes in charge of the cleavage of the components that make up the ECM, which are involved in different physiological and pathological processes that occur in living tissues. According to their function within tissues, they can be classified as: Collagenases, comprising MMP-1, MMP-8, MMP-13 and MMP-18; which can degrade interstitial collagen I, II and III, resulting in denatured collagen or gelatin. MMP-1, which is synthesized by macrophages, fibroblasts and dendritic cells, is in charge of promoting cell survival. MMP-8, which is synthesized by neutrophils, shows antitumor and anti-invasive properties due to its role in hormone receptor regulation. MMP-13, in turn, is only secreted by fibroblasts.
Gelatinase A (MMP-2) and B (MMP-9) are two members of this group shown to, among other things, degrade the type IV collagen of the basement membrane (5). MMP-2, which has a molecular weight of 72 kDa, also digests collagen I, II and III, and, under normal conditions, is expressed by the stromal cells of most tissues (macrophages, mast cells, fibroblasts, dendritic, endothelial and hematopoietic cells). MMP-9, with a molecular weight of 92 kDa, is nearly absent in normal tissues and is secreted by dendritic and hematopoietic cells, neutrophils, macrophages, mast cells, fibroblasts y lymphocytes (6).
Stromelysins, which are MMP-3, MMP-10 and MMP-11, digest several components of the ECM and the basement membrane. MMP-3 indirectly controls cell migration and is secreted by lymphocytes, endothelial cells, dendritic cells and fibroblasts. MMP-11, which is connected to adipogenesis processes, is expressed in adipose tissue in the immediate surroundings of a tumor process, is a negative regulator of pre-adipocyte differentiation and reverts mature adipocyte differentiation, which leads to the accumulation of peritumoral fibroblast-like cells, which favors tumor progression (7).
In turn, matrilysins MMP-7 and MMP-26 are structurally the simplest because they lack a hemopexin domain, which plays a role in the binding to MMP-specific substrates and endogenous inhibitors, act on cell surface molecules and are specifically expressed by tumor cells of epithelial origin. MMP-7 is secreted by osteoclasts, macrophages and endothelial cells, and is involved in angiogenesis, cell invasion and inflammation processes.
Membrane-type matrix metalloproteases (MT-MMP) are part of basement membranes and play a role in the proteolytic activity of other MMPs. These can be divided into two kinds: - Transmembrane proteins: bound to it through a hydrophobic region, MMP-14, MMP-15, MMP-16, MMP-24 - Proteins with glycophosphatidylinositol (GPI): MMP-17 and MMP-25. The most studied of these is MMP-14, which is secreted by macrophages, fibroblasts, endothelial and hematopoietic cells, and is involved in the processes of angiogenesis, adipogenesis and cell growth.
Aside from the MMPs which are grouped in families according to their function and structural similarity, some are in a class of their own: MMP-12, a metalloelastase that is mainly expressed by macrophages and affects their migratory capacity. Elastin is its main substrate. MMP-20 or enamelysin, digests the members of the amelogenin family, extracellular matrix proteins. MMP-22, whose function remains unknown. MMP-23, predominantly expressed in reproductive tissues and lacks a hemopexin domain. MMP-28, expressed in keratinocytes, plays a role in hemostasis and wound healing (6).
These functions are especially regulated by TIMPs, which can be divided into three kinds. TIMP-1 potentially inhibits the activity of most MMPs, with the exception of MT1-MMP and MMP-2, and is synthesized by neutrophils, lymphocytes, mast cells (7). TIMP-2 also inhibits the activity of most MMPs, except MMP-9, and like TIMP-1 it is synthesized by endothelial, dendritic and hematopoietic cells. TIMP-3 inhibits the activity of MMP-1, -2, -3, -9, and of MT1-MMP (5). All of them are synthesized by tissue macrophages and fibroblasts.
The connection between metalloproteinases and odontogenesis
There is evidence of the involvement of MMPs in the process of odontogenesis. The loss of proteins in the enamel matrix is known to occur during the maturation stage of the process, before tooth eruption. It has been suggested that MMP-1, -2, -3 and -9 proteins contribute to early tooth morphogenesis. In turn, TIMP-2 and the gelatinases A (MMP-2) and B (MMP-9) are co-expressed during tooth morphogenesis. The balance between MMPs and TIMPs in the pericellular environment will determine the most significant proteolytic in tissue remodeling.
MMP-9 is involved in bone resorption and the degradation of the basement membrane of the reduced enamel epithelium during tooth development, and of the ECM, associated with tooth eruption (8). Several studies indicate that MMP-9 binds to amelogenin or both are co-expressed in ameloblasts in the development of the dental germ. Their spatial distribution was co-localized in preameloblasts, secretory ameloblasts, enamel matrix and odontoblasts (9).
MMPs can promote cell movement through the hydrolysis of cadherins, which are intercellular binding proteins. The extracellular domain of E-cadherin is cleaved by MMP-3, -7 and -9, promoting cell migration, invasion and proliferation. MMP-2, MMP-3, MMP-9 and MMP-8 (10), in turn, can cleave several components of the tissular junction (TJ) complex, which favors these processes. Thus, if ameloblasts were to maintain the tight cell-cell attachment due to MMP inactivation, this would result in dysplasia during the enamel maturation stage (amelogenesis imperfecta) (11) in the absence of cell motility.
Therefore, we consider the functions of the MMPs in the vast majority of the phases and stages of tooth development (Fig. 1).
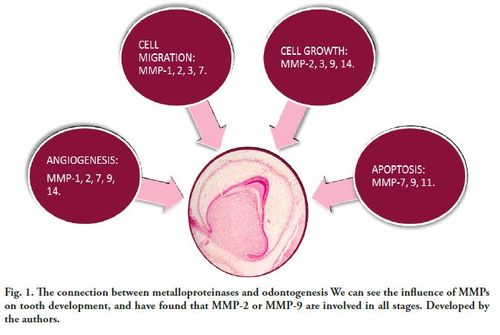
The connection between metalloproteinases and restorative dentistry
Functional polymorphisms in MMP and TIMP genes can lead to an increase or decrease in their activity, which would affect enamel formation and the development of caries. The development of dental caries is linked to MMP-2 and -20, which can cleave amelogenin, the major structural protein component of the enamel matrix (1). MMP-2 is not detected in healthy dentinal tubules, but it is detected throughout the extension of the carious lesion affecting said structures. Odontoblasts are presumed to be responsible for the secretion of MMP-2 in response to a carious lesion (12).
Upon studying the materials used in dental restoration, it was found that the hybrid layer formed by contemporary adhesives is unstable in aqueous media due to the hydrolytic degradation of both the resin adhesive and the collagen fibers. It has been shown that the endogenous proteases of the dentin organic matrix can degrade the exposed collagen fibers together with the hybrid layers if they are not protected with an adhesive monomer. MMPs are deemed to be the cause of said degradation, as that dentin contains MMP-8, MMP-2, MMP-9, MMP-3, MMP-20 (these are covered by apatite nanocrystals during the mineralization of dentin, which makes them immobile and inactive). When dentin is partially demineralized (whether by a caries process or by an acid attack caused by dental materials, lowering pH levels, creating a favorable environment), the latent forms of MMP-2 and MMP-9 are activated (13). The application of synthetic MMP inhibitors on the dentin surface after acid etching or its addition to the adhesive system promotes the integrity and stability of the restoration over time. The application of 2 % and 0.2 % chlorhexidine has been suggested for this purpose (14).
The connection between metalloproteinases and endodontics
Apical lesions, caused after pulp necrosis, mark an oxidative imbalance coupled with an increase in the activity of MMP-2 and MMP-9, which could contribute to the progression of apical periodontitis (AP). This is also seen in the gingival crevicular fluid of teeth with PA, and is resolved to normal levels with an adequate endodontic treatment (2). Variations in the levels of MMP-2, -3 and -9 are associated with untreated periapical lesions, but root canal therapy has been found to reduce their expression. A lack of a positive response to the endodontic treatment is due to an infection in the root canal system, where apical lesions remain; microbial components then induce the production of proinflammatory cytokines, playing a key role in the destruction of tissue through MMPs (15).
The connection between metalloproteinases and periodontics
Certain proteases produced by Treponema denticola, regarded as a periodontal pathogen of Socransky’s red complex, are in charge of activating pro-MMP-2, secreted in their inactive form by cells in the periodontal ligament. MMP-2 will cause the fragmentation of fibronectin, inducing apoptosis or a suppression of osteoblast differentiation, thus giving the process a destructive phenotype.
Several studies have suggested that Porphyromonas gingivalis could also activate MMP-2 (16) through its gingipains, also increasing the migration of monocytes by increasing the activation and expression of MMP-9 (17), which indirectly causes tissue lesions.
Since studies have shown an increased expression and activity of MMP-9 in patients with periodontitis, in comparison with MMP-2, it could be considered a predictive indicator of the disease activity (3). A decrease in the levels of MMP-1, -8, -9, -12 and -13 in the gingival crevicular fluid has been observed after treating localized aggressive periodontitis with scaling and planing, and with systemic antibiotic therapy, which establishes a correlation with healthy periodontal tissues. Therefore, it can be deduced that the presence of said proteases is detrimental to the periodontium since it favors a destructive microenvironment (18, 19).
While reviewing the situation of peri-implant tissue, we found that a connection has been established between the presence of MMP-1, -8 and -13 in peri-implant sulcular fluid and active periods of annual vertical bone loss. In these cases, MMP-8 is considered as a possible marker for progressive bone loss in implants (20).
Upon studying patients undergoing orthodontic treatment, we found that the application of orthodontic forces to a tooth generates tension and compression sites in the periodontal ligament, which affects remodeling of the alveolar bone and gingival tissue. MMP-1, -8, -2 and -9 have been proven to be in higher levels in compression and tension sites. Chemical inhibition of MMP-9 reduces orthodontic dental movements (4). Gingival hypertrophy without signs of inflammation can be caused by a reaction of the gingival tissue to the mechanical stress induced by orthodontic forces, triggering MMP-9 activity, which makes it a marker in gingival crevicular fluid and gingival tissue for this situation. MMP-9 levels are higher in case of chronic gingivitis, but lower than in the presence of active periodontal disease (21).
The connection between metalloproteinases and tumors
The presence of these proteases in tooth development suggests that they participate in the embryonic development of the dental organ, which is why various authors have studied their expression in odontogenic tumors, as well as in other dysplasias and pathological processes (5, 6).
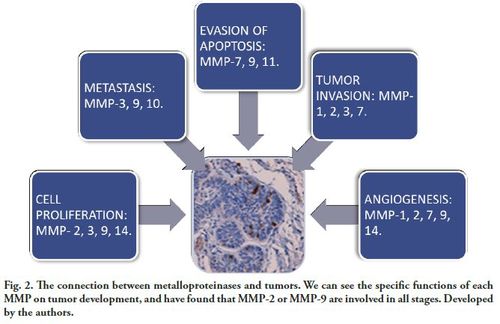
MMPs are the main mediators in alterations observed in the tumor microenvironment during cancer progression. They have a fundamental role in tumor growth, through the degradation of connective tissue and the basement membrane components, and also through the activation of growth factors, cell surface receptors for adhesion molecules and chemokines. This interaction alters the cellular response, which in turn makes tumor cells less adhesive and, therefore, more likely to migrate and result in metastases. MMPs can play different roles during cancer progression, depending on the tumor stage (Fig. 2).
In early tumor stages, the MMP-3 and MMP-7-mediated proteolysis of specific proteins that bind growth factors contributes to cell proliferation. MMP-7, in turn, causes the cleavage of FAS (apoptosis antigen 1) receptor ligands, thus suspending cellular apoptotic activity (7). MMP-7 indirectly modulates the activity of diverse molecules by cleaving syndecan-1 (transmembrane protein that contributes to tumor suppression) from the cell surface. This MMP is expressed by osteoclasts at the tumor-bone interface, inducing osteolysis and the resulting bone metastasis (7).
Afterwards, the cleavage of E-cadherin and CD44 adhesion molecules by MMP-3 activates tumor cell motility, and facilitates the formation of metastases. Cell migration and invasion are very much linked to the presence of MMP-14 in the cell membrane, and to the binding of MMP-2 to mesenchymal cell integrins (22).
The differentiation and accumulation of peritumoral adipose tissue favors the progression and survival of cancer cells. MMP-14 contributes to the coordination of adipocyte differentiation. On the other hand, MMP-11, which is expressed in adipose tissue during tumor invasion, negatively regulates adipogenesis, thus promoting tumor progression (7).
The angiogenesis process, which involves the formation of new blood vessels from pre-existing ones, enables tumor growth and is essential for the dissemination of tumor cells. Most MMPs are involved in tumor angiogenesis, mostly MMP-2, -9, and -14, and to a lesser extent MMP-1 and -7. MMP-9 is in charge of regulating the bioavailability of vascular endothelial growth factor (VEGF), a potent inducer of tumor angiogenesis, and is also connected to vasculogenesis (7).
Lymphatic invasion and lymph node metastases are associated with an increased expression of MMP-1, -2 and -3 (7).
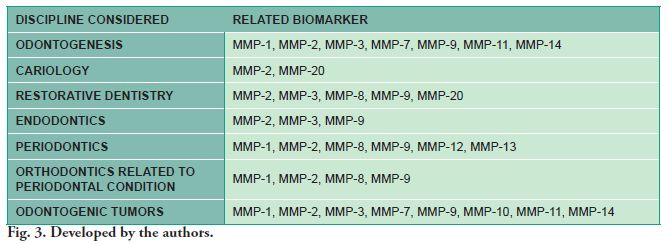
Fig. 3 summarizes the connection between the disciplines considered and the MMP that have a more prominent role. MMP-2 or MMP-9 were found to be involved in all of the areas considered in the study.
Conclusion
It is concluded that MMP-2 and MMP-9 are involved in all of the pathological processes mentioned above, both during the development of the dental germ and through their influence on the proliferation and invasion of odontogenic tumors, their participation in the degradation of dental and periodontal tissues by promoting the formation of a carious lesion, gingivitis and periodontitis; confirming that this family of proteins can play a major role in various oral biological processes. Moreover, this means that MMPs are currently considered significant biomarkers for research in the different areas of specialization of dentistry.
Referencias
2. Dezerega A, Madrid S, Mundi V, Valenzuela MA, Garrido M, Paredes R, García-Sesnich J, Ortega AV, Gamonal J, Hernández M. Pro-oxidant status and matrix metalloproteinases in apical lesions and gingival crevicular fluid as potential biomarkers for asymptomatic apical periodontitis and endodontic treatment response. J Inflamm (Lond). 2012; 9(1): 8.
3. Kim KA, Chung SB, Hawng EY, Noh SH, Song KH, Kim HH, Kim CH, Park YG. Correlation of expression and activity of matrix metalloproteinase-9 and -2 in human gingival cells of periodontitis patients. J Periodontal Implant Sci. 2013; 43(1): 24-29.
4. Grant M, Wilson J, Rock P, Chapple I. Induction of cytokines, MMPs9, TIMPs, RANKL and OPG during orthodontic tooth movement. Eur J Orthod. 2013; 35(5): 644-651.
5. Cascales M, Álvarez-Gómez JA. Metaloproteinasas, matriz extracelular y cáncer. An R Acad Nac Farm, 2010, 76 (1): 59-84.
6. Coronato S, Laguens G, Vanda Di Girolamo V. Rol de las metaloproteinasas y sus inhibidores en patología tumoral. Medicina (Buenos Aires) [en línea] 2012; 72: 495-502 citado 11 de agosto 2016. Disponible en http://www.scielo.org.ar/pdf/medba/v72n6/v72n6a12.pdf.
7. Kessenbrock K, Plaks V, Werb Z. Matrix Metalloproteinases: Regulators of the Tumor Microenvironment. Cell. 2010; 141(1): 52-67.
8. Basi DL, Hughes PJ, Thumbigere-Math V, Sabino M, Mariash A, Lunos SA, Jensen E, Gopalakrishnan R. Matrix metalloproteinase-9 expression in alveolar extraction sockets of zoledronic acid-treated rats. J Oral Maxillofac Surg. 2011; 69:2698–2707.
9. Feng J, McDaniel JS., Chuang HH., Huang O, Rakian A., Xu X., Steffensen B., Donly KJ., MacDougall M, Chen S. Binding of amelogenin to MMPs-9 and their co-expression in developing mouse teeth. J Mol Hist. 2012; 43(5): 473–485.
10. Schubert-Unkmeir A, Konrad C, Slanina H, Czapek F, Hebling S, Frosch M. Neisseria meningitidis induces brain microvascular endothelial cell detachment from the matrix and cleavage of occludin: a role for MMPs-8. PLoS Pathog [en línea]. 2010; 29; 6(4):e1000874 citado: 11 de agosto 2016. Disponible en: http://dx.doi.org/10.1371/journal.ppat.1000874
11. Bartlett JD, Smith CE. Modulation of cell-cell Junctional complexes by matrix metalloproteinases. J Dent Res. 2013; 92(1): 10-17.
12. Boushell LW, Nagaoka H, Nagaoka H, Yamauchi M. Increased Matrix Metalloproteinase-2 and Bone Sialoprotein Response to Human Coronal Caries. Caries Res. 2011; 45(5): 453-459.
13. Mazzoni A, Scaffa P, Carrilho M, Tjäderhane L, Di Lenarda R, Polimeni A, Tezvergil-Mutluay A, Tay FR, Pashley DH, Breschi L. Effects of Etch-and-Rinse and Self-etch Adhesives on Dentin MMPs-2 and MMPs-9. J Dent Res. 2013; 92(1): 82-86.
14. Montagner AF, Sarkis-Onofre R, Pereira-Cenci T, Cenci MS. MMPs Inhibitors on Dentin Stability: A Systematic Review and Meta-analysis. J Dent Res. 2014; 93(8): 733-743.
15. Menezes-Silva R, Khaliq S, Deeley K, Letra A, Vieira AR. Genetic Susceptibility to Periapical Disease: Conditional Contribution of MMPs2 and MMPs3 Genes to the Development of Periapical Lesions and Healing Response. J Endod. 2012; 38(5): 604-607.
16. Miao D, Godovikova V, Qian X, Seshadrinathan S, Kapila YL, Fenno JC. Treponema denticola upregulates MMPs-2 activation in periodontal ligament cells: interplay between epigenetics and periodontal infection. Arch Oral Biol. 2014; 59(10): 1056-1064.
17. Zhou J, Zhang J, Chao J. Porphyromonas gingivalis Promotes Monocyte Migration by Activating MMPs-9. J Periodontal Res. 2012; 47(2): 236-242.
18. Gonçalves PF, Huang H, McAninley S, Alfant B, Harrison P, Aukhil I, Walker C, Shaddox LM. Periodontal Treatment Reduces Matrix Metalloproteinase Levels in Localized Aggressive Periodontitis. J Periodontol. 2013; 84(12): 1801-1808.
19. Şurlin P, Oprea B, Solomon SM, Popa SG, Moţa M, Mateescu GO, Rauten AM, Popescu DM, Dragomir LP, Puiu I, Bogdan M, Popescu MR. Matrix metalloproteinase -7, -8, -9 and -13 in gingival tissue of patients with type 1 diabetes and periodontitis. Rom J Morphol Embryol. 2014; 55(3 Suppl): 1137-1141.
20. Arakawa H, Uehara J, Hara ES, Sonoyama W, Kimura A, Kanyama M, Matsuka Y, Kuboki T. Matrix metalloproteinase-8 is the major potential collagenase in active periimplantitis J Prosthodont Res. 2012; 56(4): 249-255.
21. Şurlin P, Rauten AM, Pirici D, Oprea B, Mogoantă L, Camen A. Collagen IV and MMPs-9 expression in hypertrophic gingiva during orthodontic treatment. Rom J Morphol Embryol. 2012; 53(1): 161-165
22. Wolf K, Friedl P. Mapping proteolytic cancer cell-extracellular matrix interfaces. Clin. Exp. Metastasis 2009; 26(4): 289-298.
23. Paula-Silva FW, da Silva LA, Kapila YL. Matrix metalloproteinase expression in teeth with apical periodontitis is differentially modulated by the modality of root canal treatment. J Endod. 2010; 36(2): 231-237.
24. Zehnder M, Wegehaupt FJ, Attin T. A first study on the usefulness of matrix metalloproteinase 9 from dentinal fluid to indicate pulp inflammation. J Endod. 2011; 37(1): 17-20.
25. Capelli J Jr, Kantarci A, Haffajee A, Teles RP, Fidel R Jr, Figueredo CM. Matrix metalloproteinases and chemokines in the gingival crevicular fluid during orthodontic tooth movement. Eur J Orthod. 2011; 33(6): 705-711.
26. Chen X, Li N, LeleYang, Liu J, Chen J, Liu H. Expression of collagen I, collagen III and MMPs-1 on the tension side of distracted tooth using periodontal ligament distraction osteogenesis in beagle dogs. Arch Oral Biol. 2014; 59(11): 1217-1225.
27. Canavarro C, Teles RP, Capelli Júnior J. Matrix metalloproteinases -1, -2, -3, -7, -8, -12, and -13 in gingival crevicular fluid during orthodontic tooth movement: a longitudinal randomized split-mouth study. Eur J Orthod. 2013; 35(5): 652-658.
28. Díaz Caballero A, Méndez Cuadro D, Martínez Serrano E, Orozco Páez J, Velásquez M. Metaloproteinasas de la matriz en Odontología y sus consideraciones desde el campo de la química computacional. Rev Cubana Estomatol [en línea]. 2014: 51(1):80-92 citado: 11 de agosto 2016. Disponible en: http://scielo.sld.cu/pdf/est/v51n1/est09114.pdf
29. Rai B, Kaur J, Jain R, Anand SC. Levels of gingival crevicular metalloproteinases 8 and -9 in periodontitis. Saudi Dent J [en línea] 2010; 22(3): 129-131 citado: 11 de agosto 2016. Disponible en: http://www.saudidentaljournal.com/article/S1013-9052(10)00044-1/pdf

