











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo

 Permalink
Permalink
Introducción
El complejo respiratorio bovino (CRB) y la diarrea neonatal (DN) son dos de las causas más significativas de morbilidad y mortalidad en terneros de lechería especializada (Cantor et al., 2021). Los virus de la diarrea viral bovina (VDVB) y herpesvirus bovino 1 (HVB-1) son patógenos importantes asociados con la presentación del CRB (Chamorro et al., 2014) y el VDVB está también asociado con la presentación de DN (Caffarena et al., 2021). Uno de los factores más críticos que afecta la resistencia de los terneros neonatales frente a las infecciones es el manejo del calostro (Lorenz et al., 2021). En lecherías especializadas se han establecido valores aceptables para el manejo de la transferencia pasiva (TP), un buen indicador es cuando se registra un máximo de 20 % de los terneros con TP aceptable y un 10 % con fallo de la TP a nivel de hato (de Souza et al., 2021; Lombard et al., 2020). No obstante, es importante señalar que el virus de la leucosis bovina enzoótica (VLBE) es uno de los patógenos más importantes asociado con disfunción del sistema inmune en ganado adulto, lo que puede alterar la calidad de la calostrogénesis (Frie et al., 2016); así mismo, por el efecto inmunosupresor del VDVB y HVB-1 (Segura-Correa et al., 2016).
Por otro lado, si el CRB y la DN afectan al 33 % de los terneros antes del destete (Cantor et al., 2021) y el riesgo de infección con el VLBE es más alto durante el periodo neonatal (Evermann, 2014), el incremento de la TP de baja calidad reducirá la capacidad de los terneros para sobrevivir y responder de forma satisfactoria a los estímulos antigénicos (Windeyer et al., 2012), aumentará la prevalencia del VLBE (Evermann, 2014), los tratamientos antimicrobianos y el uso de antiinflamatorios para controlar las infecciones asociadas con neumonía y DN (Cantor et al., 2021; Krehbiel, 2020). Si bien los antibióticos se han utilizado en la producción de alimentos para animales como promotores del crecimiento y la salud, su uso representa un grave peligro para la salud pública veterinaria porque favorece la emergencia de cepas bacterianas con resistencia múltiple a los antibióticos (Ma et al., 2021; Van et al., 2020; Vlasova y Saif, 2021). Por tanto, es una necesidad prioritaria de la industria ganadera desarrollar alternativas que reduzcan la dependencia a los antibióticos y que apoyen los programas de prevención y control de las enfermedades infecciosas.
En ese sentido, se ha tratado de mejorar la resistencia de las terneras frente a las infecciones más comunes en el hato y evitar la transmisión de agentes patógenos mediante el suministro de calostro pasteurizado seropositivo frente al VLBE, VDVB y HVB-1 para reducir el riesgo de infección (Gröner et al., 2018; Pitino et al., 2021; Sandoval-Monzón et al., 2021) o bien mediante el suministro de reemplazador de calostro (RC) (Chamorro et al., 2014; Lomónaco et al., 2020). Asimismo, mediante estrategias nutricionales que favorecen el desarrollo y la maduración del tracto gastrointestinal y del sistema inmune (Ballou et al., 2019; Vlasova y Saif, 2021), Vega et al. (2020), por ejemplo, demostraron que el uso de suplemento de inmunidad pasiva a base de IgY en la leche durante las primeras dos semanas de vida redujo la duración y la gravedad de la diarrea; Kargar et al. (2020) determinaron que los terneros suplementados de forma extendida con calostro durante 14 días fueron más resistentes para enfermar de diarrea y neumonía, y Chamorro et al. (2017) suplementaron los terneros con RC por 14 días para reducir la incidencia de diarrea y el uso de antibióticos antes del destete. Asimismo, el uso de pre y probióticos como suplemento alimenticio ha generado resultados contrapuestos, no obstante, los efectos beneficiosos se observan en el crecimiento y durante el tratamiento de alguna enfermedad por el efecto modulador sobre la respuesta inmune y la salud gastrointestinal (Cangiano et al., 2020; Vlasova y Saif, 2021; Zamojska et al., 2021).
El manejo combinado del plan nutricional con algunas de las estrategias antes mencionadas genera los mejores resultados para reducir el impacto del CRB y la DN en los reemplazos (Krehbiel, 2020; Lorenz et al., 2021; Vlasova y Saif, 2021); sin embargo, el uso de suplementos y la alimentación extendida con calostro es costosa (Cantor et al., 2021), lo que sugiere la necesidad de desarrollar alternativas que ayuden a fortalecer el efecto de la nutrición sobre el estado de salud a menor costo. Por esto, la lactoterapia es una estrategia promisoria y barata para activar la respuesta inmune de forma activa e inespecífica, en especial, como coadyuvante en el uso de antibióticos y la vacunación (Bielecka et al. 2022; Prieto-Cristancho, 2012). En efecto, es necesario determinar si las terneras calostradas con calostro o RC son inmunocompetentes frente al VLBE, VDVB y HVB-1 y si el efecto de la transferencia pasiva y la lactoterapia puede mejorar la inmunidad humoral contra estos virus. Por lo tanto, los objetivos de esta investigación fueron evaluar la inmunocompetencia frente al VLBE, VDVB y HVB-1 asociada con el suministro de calostro materno y RC en terneras desde el nacimiento así como valorar el efecto de la transferencia pasiva y la lactoterapia sobre la inmunocompetencia de las terneras desde los 55 hasta los 102 días de edad.
Materiales y métodos
Manejo de animales y diseño experimental
Este estudio se desarrolló en una lechería comercial ubicada en Alajuela, Costa Rica. Los procedimientos sobre manejo de los animales fueron revisados y aprobados por el Comité Institucional para el Cuidado y Uso de los Animales de la Universidad de Costa Rica (Oficio CICUA-067-2020). La lechería mostró altos porcentajes de vacas seropositivas para el VLBE (95,1 % (78/82)), VDVB (29,3 % (17/58)) y HVB-1 (95,7 % (45/47)) (Pichardo-Matamoros et al., 2020). 40 terneras, recién nacidas, de diferentes razas lecheras (jersey, holstein, pardo suizo, ayrshire, brahman, gyr y sus cruces) fueron asignadas a uno de dos grupos de forma sistemática desde julio hasta diciembre, formando bloques con base en la fecha de nacimiento y el peso, como sigue: (1) suministro de CM a una dosis media de ≈ 171 g de IgG en 1,8 L (grupo CM o A1, n=20) y (2) suministro de RC (Premolac® 150, Zinpro Corporation, Eden Prairie, MN) a una dosis media de ≈ 150 g de IgG en 1,5 L (grupo RC o A2, n=20). Las terneras fueron separadas de su madre entre 20-45 min después del parto para evitar la succión de calostro, se alojaron en jaulas de 1,5 m de largo × 1 m de ancho de forma individual o en pareja y se les curó el ombligo con solución de alcohol-yodado al 70 %. Mayores detalles sobre los terneros se describen en la literatura (Pichardo-Matamoros et al., 2023). Se recolectó en la finca el CM de 30 vacas donadoras, aparentemente sanas y sin mastitis; una vez reunidos 103,2 L de calostro, se homogeneizaron y formularon dosis usando recipientes nuevos, las cuales se pasteurizaron a 60 °C × 30 min y conservaron a -45 °C hasta su uso. La mezcla de CM contenía 13,2 % de proteína cruda y 6 % de grasa sobre base de materia seca, recuento total de 80 UFC/mL, coliformes totales < 10 UFC/mL, E. coli < 10 UFC/mL, resultó negativa para Mycobacterium avium ssp. Paratuberculosis y seropositiva para el VLBE, VDVB y HVB-1.
El calostro fue suministrado entre 2,4 y 3,6 horas después del nacimiento empleando chupón. La segunda alimentación, 12 horas posteriores a la ingesta de calostro, consistió en una dosis de leche pasteurizada similar a la ofrecida durante el calostrado. Luego, las terneras fueron trasladadas y alojadas en un corral grupal para recién nacidos de 8,3 m de largo × 2,4 m de ancho. A partir de la tercera alimentación, las terneras fueron alimentadas con 2 L de leche fresca cada 12 horas (0400 y 1600 horas) empleando chupón o cubeta. A los 14 días de edad, las terneras fueron trasladadas de forma sistemática y alojadas en un establo de 6,6 m de largo × 5,9 m de ancho donde se les ofreció heno (4,8 % de proteína cruda y 1,8 % de grasa sobre base de materia seca), concentrado iniciador (20 % de proteína cruda y 2,5 % de grasa sobre base de materia seca) y agua potable de forma restringida hasta el destete.
El destete se llevó a cabo alrededor de los 56 días de edad. Una vez ocurrido el destete, las terneras fueron trasladadas de forma sistemática a otro establo de 7,9 m de largo × 2,3 m de ancho, con piso de plástico a una altura de 30 cm desde el suelo. En esta etapa posdestete el programa de alimentación consistió en alimento balanceado (13,7 % de proteína cruda y 2,5 % de grasa sobre base de materia seca), forraje (14,8 % de proteína cruda y 2 % de grasa sobre base de materia seca) y agua potable ad libitum hasta el final del estudio. En general, los establos tenían techo, paredes y suelo de concreto y cama de aserrín, la cual se cambió cada tres días.
Alrededor de los 55 días de edad, se inició el ensayo clínico con CM intramuscular, el cual se desarrolló de forma sistemática en tres momentos y de forma paralela en réplicas pares dentro de un mismo grupo de calostrado para formar dos grupos, como sigue: (1) no aplicación intramuscular de CM (grupo N0, n=17) y (2) aplicación intramuscular de 5 mL de CM, tres dosis cada cinco días (grupo N1, n=18) formando grupos no balanceados debido a la ocurrencia de algunas muertes en terneras menores a los 55 días de edad; así como, cuatro combinaciones entre los niveles de A × N (A1N0, n=10; A1N1, n=11; A2N0, n=7 y A2N1, n=7). El calostro fue obtenido desde una vaca jersey seropositiva frente al VLBE, VDVB y HVB-1, de diez años de edad, BCS=3,50 y lactancia número ocho. El calostro fue pasteurizado a 60 °C × 30 min y contenía medias de 18,7 g/100g de proteína, 9,98 g/100 g de caseína, 4,43 g/100 g de grasa, 1,68 g/100 g de lactosa, 264 µg/100 g de vitamina A, 100 g/L de IgG, recuento total de 20 UFC/mL, coliformes totales < 10 UFC/mL, E. coli < 10 UFC/mL, resultó negativo para la presencia de Mycobacterium avium ssp. Paratuberculosis y seropositivo para el VLBE, VDVB y HVB-1.
Toma y procesamiento de muestras
Muestras de sangre fueron recolectadas por punción de la vena coccígea en vacas y desde la vena yugular en terneras empleando el sistema Vacutainer® y tubos de ensayo con y sin anticoagulante debidamente rotulados. El suero fue separado de la sangre a 3500 g × 5 min. Alícuotas de 1,5 mL de sangre completa y suero fueron depositadas en microtubos Eppendorf de 1,5 mL, las cuales fueron conservadas a -45 °C hasta su uso. En general, las muestras fueron recolectadas antes y después del calostrado, a los 20 y 45 días de edad en los grupos CM y RC; luego, a los 55 días de edad durante la aplicación de la primera dosis de CM intramuscular, a los 60 días de edad cuando se aplicó la segunda dosis de CM y primera dosis de vacuna, a los 65 días de edad cuando se aplicó la tercera dosis de CM, a los 81 días de edad cuando se aplicó la segunda dosis de vacuna y a los 102 días cuando finalizó el ensayo clínico con CM intramuscular. Adicionalmente, muestras de sangre, heces o secreción respiratoria de terneras enfermas fueron recolectadas y conservadas a -45 °C hasta su uso.
Valoración de la inmunocompetencia
La inmunocompetencia de las terneras fue valorada de forma general mediante el estatus de salud y de forma específica mediante la vigilancia activa de la exposición a los virus y la inmunización con Express® FP 10-HS (Boehringer Ingelheim Vetmedica, Germany) a base de virus vivo modificado de HVB-1 (Cepa Colorado), VDVB-1 (cepa Singer citopática), VDVB-2 (cepa 296 citopática), virus parainfluenza tipo 3 y virus respiratorio sincitial bovino; así como bacterinas de Histophilus somni, Leptospira canicola, L. grippotyphosa, L. hardjo, L. icterohaemorrhagiae y L. Pomona, la cual fue administrada según instrucciones del fabricante.
Estatus de salud
Los casos de DN y trastornos respiratorios fueron registrados diariamente. Los casos de diarrea fueron registrados cuando las terneras defecaron desde heces blandas hasta fluidas, con aumento de la frecuencia y pérdida del control fisiológico de la evacuación de las heces; así como la presentación de deshidratación, pérdida de peso, quejidos, dolor abdominal, decúbito o muerte. Mientras tanto, los eventos asociados con enfermedad respiratoria fueron registrados cuando la ternera mostró secreción patológica de las fosas nasales, tos, apatía, disnea, fiebre, decúbito o muerte (Karle et al., 2019; Kim et al., 2020; Timmerman et al., 2005).
Pruebas serológicas
Muestras de suero fueron analizadas para determinar la concentración de anticuerpos frente al VLBE, VDVB y HVB-1 empleando pruebas de inmunoabsorción ligada a enzimas. En vacas y terneras, la presencia de anticuerpos frente al VLBE, VDVB y HVB-1 fueron determinados usando los reactivos ID Screen® BLV Competition, ID Screen® BVD p80 Antibody Competition e ID Screen® IBR gB Competition (ID.vet, Grabels, France) según instrucciones del fabricante, respectivamente. el estatus sanitario y los niveles de anticuerpos frente a los virus fueron evaluados mediante el porcentaje de inhibición (PI), el cual fue determinado mediante la fórmula propuesta por Machado et al. (2015): PI (%) = ((DO Control negativo - DO Muestra)/DO Control negativo) × 100.
Además, la respuesta humoral frente a la inmunización en terneras contra el VDVB fue determinada mediante la prueba de virus neutralización (PVN). Los sueros fueron probados para detectar anticuerpos en contra de la cepa citopática NADL-1 utilizada a una concentración de 100 DICT50. Primeramente, los sueros fueron inactivados a 56 °C × 30 min y se realizaron diluciones seriadas desde 1:2 hasta 1:4096 del suero en 50 μL de medio de cultivo; luego, se inocularon dos pocillos por cada dilución en una placa de microtitulación de poliestireno de 96 pocillos (ThermoFisher Scientific, USA), se incubó la placa durante 72 horas y se evaluó el efecto citopático en células de riñón de bovino (MDBK, Madin-Darby Bovine Kidney) según lo descrito en la literatura (Chamorro et al., 2014; Klimowicz-Bodys et al., 2021). Los microtítulos de anticuerpos frente al VDVB fueron transformados empleando la función logarítmica. El estatus seronegativo frente al VDVB fue definido como un título de anticuerpos séricos inferior a 2, igual a un título log2 de 0; así mismo, por un PI < 60.
Detección del VLBE, VDVB y HVB-1 mediante PCR (Polymerase Chain Reaction)
La extracción del ARN viral se realizó a partir de 250 µL de suero empleando Trizol (Ambion, USA). El ARN extraído fue utilizado para sintetizar ADN complementario empleando RevertAid H Minus First Strand cDNA Synthesis Kit (Fermentas, Lituania). El ADN genómico fue aislado a partir de 300 µL de sangre con EDTA usando Wizard® Genomic DNA Purification Kit (Promega Corporation, Madison, USA). Todos los procedimientos fueron realizados según instrucciones del fabricante. El ARN, ADN complementario y ADN genómico obtenidos fueron conservados a -75 °C hasta su uso. Cada reacción de PCR fue realizada empleando cebadores específicos para cada virus (tabla 1).
Tabla 1: Cebadores utilizados en el estudio

1Cebadores Env1/Env3 y Env2/Env3 amplifican la región gp51 del VLBE. Los cebadores TS1/P2, TS2/P2 y TS3/P2 amplifican la región Erns del virus de la enfermedad de las fronteras, VDVB-2 y VDVB-1, respectivamente. Mientras, los cebadores BHV1-IgE-F/BHV1-IgE-R amplifican la glicoproteína E de herpesvirus bovino 1.
El VLBE fue identificado mediante PCR anidado. El volumen final de cada reacción fue de 20 µL y consistió en 10 µL de Phusion U Multiplex PCR Master Mix #F562S (Invitrogen, USA), 1,2 µL de cada cebador Env1(+)/Env3(-) o Env1(+)/Env4(-) en la primera ronda y Env2(+)/Env3(-) (0,3 µM/µL de cada cebador) en la segunda ronda, 1 µL de ADN y H2O libre de ARNasas (Invitrogen, USA) para completar el volumen. Las condiciones de la reacción fueron las siguientes: desnaturalización inicial a 98 °C × 3 min, 35 ciclos de desnaturalización a 98 °C × 20 s, anillamiento a 70 °C × 30 s y extensión a 72 °C × 15 s, seguido de una extensión final a 72 °C × 5 min. Se usó ADN de la cepa 280918-4418-CR aislada en Costa Rica como control positivo (número de acceso al GenBank: MN830810). El producto esperado fue de 725 (Env1/Env3) o 912 pb (Env1/Env4) en la primera ronda y 443 pb (Env2/Env3) en la segunda ronda (Modificado de, Khudhair et al., 2016).
La presencia del VDVB fue determinada empleando los cebadores TS1(+), TS2(+) y TS3(+) con P2(-) dirigidos a la región Erns del VDVB (Sullivan y Akkina, 1995). El volumen final de cada reacción de PCR en formato múltiple fue de 20 µL y consistió en 10 µL de Phusion U Multiplex PCR Master Mix #F562S (Invitrogen, USA), 1 µL de cada cebador TS1, TS2 y TS3 (0,25 µM/µL), 1,4 μL (0,35 μM/μL) del cebador P2, 3,6 μL de H2O libre de ARNasas (Invitrogen, USA) y 2 µL de ADN complementario. Las condiciones de la reacción fueron las siguientes: desnaturalización inicial a 98 °C × 1 min, 35 ciclos de desnaturalización a 98 °C × 15 s, anillamiento a 55 °C × 30 s y extensión a 72 °C × 15 s, seguido de una extensión final a 72 °C × 5 min. Los productos esperados fueron 566 pb (TS1/P2 para VEF), 448 pb (TS2/P2 para VDVB-2) y 223 pb (TS3/P2 para VDVB-1) (Sullivan y Akkina, 1995). Se utilizó ADN complementario de la cepa 560615-F0 aislada en Costa Rica como control positivo (número de acceso al GenBank: MT024569).
HVB-1 fue identificado mediante el PCR descrito por Wernike et al. (2011). El volumen final de cada reacción de PCR fue de 20 µL y consistió en 10 µL de Phusion U Multiplex PCR Master Mix #F562S (Invitrogen, USA), 1 µL de cada cebador IBR-IgE-F(+)/IBR-IgE(-) (0,25 µM/µL), 7 μL de H2O libre de ARNasas (Invitrogen, USA) y 1 µL de ADN. Las condiciones de la reacción fueron las siguientes: desnaturalización inicial a 98 °C × 2 min, 35 ciclos de desnaturalización a 98 °C × 20 s, anillamiento a 63 °C × 30 s y extensión a 72 °C × 15 s, seguido de una extensión final a 72 °C × 5 min. Se usó ADN de la cepa Colorado contenida en la vacuna comercial Express® FP 10-HS (Boehringer Ingelheim Vetmedica, Germany) como control positivo. El producto esperado fue de 106 pb.
Los productos amplificados por PCR fueron teñidos con GelRed Nucleic Acid Stain 10000X in Water (Biotium, USA) y separados en gel de agarosa al 2 %. Se emplearán 2 μl de buffer de muestra 6X DNA Loading Dye #R0611 (Fermentas, Lituania) y 1 μl (0,5 μg/μl) de GeneRulerTM 100 bp DNA Ladder Plus #SM0321 (Fermentas, Lituania). La separación de los amplicones se realizó usando el equipo de electroforesis Fotodyne Model n.o 1-1430 a 80v × 40 min, una vez separados se observaron en el capturador de imágenes e iluminador ultravioleta (UVP BioDoc-1tTM Imaging System/FirstLight UV illuminator, USA).
Cálculos y análisis estadístico
Primeramente, se evaluó el efecto del CM y RC sobre la TP y evolución de los niveles de anticuerpos desde el nacimiento hasta los 102 días de edad con base en el PI y luego se valoró el efecto del CM intramuscular desde los 55 hasta los 102 días de edad sobre la competencia inmune frente al VDVB. Los niveles de anticuerpos medidos por el PI para el VLBE, VDVB y HVB-1, así como los títulos de anticuerpos obtenidos por la PVN frente al VDVB, se analizaron utilizando el procedimiento mixto para medidas repetidas con el método de máxima verosimilitud restringida y empleando el criterio de información de Akaike corregido (AICc, Corrected Akaike Information Criteria) para elegir la mejor estructura de covarianza repetida. Los efectos fijos en el modelo fueron el tratamiento o estatus frente al VDVB, la edad y la interacción. Las terneras fueron incluidas como variable aleatoria. Los valores extremos fueron eliminados de los análisis. Las diferencias entre las medias de mínimos cuadrados de los distintos tratamientos se compararon utilizando la prueba HSD de Tukey-Kramer.
Los niveles de anticuerpos medidos por el PI para el VDVB fueron transformados a variable dicotómica para determinar las proporciones de animales seropositivos (PI ≥ 60 %) y seronegativos (PI ≤ 59 %) haciendo énfasis en la asociación del estatus seronegativo del grupo CM versus RC con la inmunización. Un análisis de contingencia fue realizado para determinar la asociación entre los tratamientos con la ocurrencia de enfermedad y un análisis de regresión logística fue utilizado para comparar la proporción de terneras seronegativas frente al VDVB entre grupos CM versus RC, las diferencias significativas fueron determinadas por la prueba exacta de Fischer y la prueba Ji cuadrado para la razón de máxima verosimilitud, respectivamente. Todos los análisis estadísticos fueron realizados empleado el programa estadístico JMP® Pro 15.1.0 (SAS Institute Inc., Cary, NC) y el nivel de significancia fue establecido en P < 0,05.
Resultados
Estatus sanitario
En este estudio, 72 vacas recién paridas fueron evaluadas para determinar la presencia de anticuerpos frente al VLBE, VDVB y HVB-1. Como resultado, se confirmaron altos porcentajes de vacas seropositivas para el VLBE (91,6 % (66/72)), VDVB (94,4 % (68/72)) y HVB-1 ((97,2 % (70/72)). Asimismo, se determinó el estatus sanitario de las terneras frente a los virus al nacimiento, antes del suministro de calostro. Los resultados indicaron el nacimiento de tres terneras seropositivas con infección múltiple ocasionada por el VLBE, VDVB y HVB-1, así como el nacimiento de terneras inmunocompetentes con infección transitoria y 13 terneras sospechosas de nacer con infección persistente para el VDVB (tabla 2).
Tabla 2: Caracterización de las evidencias serológicas y antigénicas frente a los virus en terneras al nacimiento
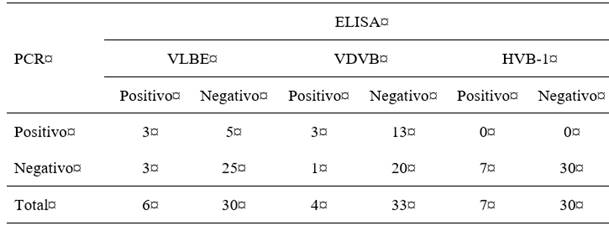
PCR=reacción en cadena de la polimerasa; ELISA=ensayo de inmunoabsorción ligado a enzimas; VLBE=virus de la leucosis bovina enzoótica; VDVB=virus de la diarrea viral bovina; HVB-1=herpesvirus bovino 1
Además, durante el ensayo se registraron 25 eventos de diarrea y 27 casos de neumonía. Como resultado, siete muestras de heces y una de secreción respiratoria resultaron positivas para la presencia del VDVB (15,3 % (8/52)), ninguna muestra fue positiva para HVB-1 en los casos de neumonía ni en los casos de mortalidad registrados (14,6% (6/41)) se detectó el VDVB y HVB-1 como etiología. Por su parte, el análisis de regresión logística mostró que la probabilidad para enfermar en el grupo CM versus el grupo RC y durante el ensayo con CM intramuscular no fue diferente, por lo que, la competencia inmune de las terneras fue independiente del tratamiento recibido (tabla 3).
Tabla 3: Análisis de asociación entre los tratamientos con la presentación de enfermedad en terneras
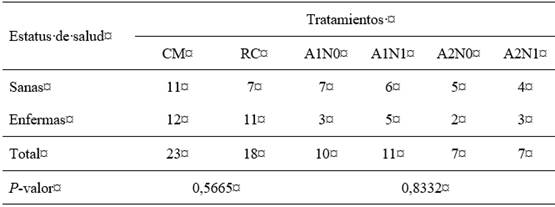
CM=grupo con suministro de calostro materno; RC=grupo con suministro de reemplazador de calostro; A1N0=grupo CM + sin aplicación de CM intramuscular; A1N1=grupo CM + aplicación de CM intramuscular; A2N0=grupo RC + sin aplicación de CM intramuscular; A2N1=grupo RC + aplicación de CM intramuscular.
Transferencia pasiva, estatus de salud e inmunocompetencia posnatal
La ingesta de CM o RC generó un aumento significativo del PI asociado con la presencia de anticuerpos frente a VLBE, VDVB y HVB-1 a los dos días de edad en comparación con el PI mostrado al nacimiento (fig. 1a), pero la diferencia entre grupos a los dos días de edad no fue significativa para el VLBE (P=0,7641) y VDVB (P=0,9315), mientras la diferencia entre las medias de los niveles de anticuerpos para HVB-1 entre el grupo CM versus RC fue significativa (P=0,0051) (tabla 4). Con respecto al VLBE, el nacimiento de terneras seronegativas y la posterior seroconversión cuando ingieren CM o RC seropositivo indicó que las terneras adquirieron protección. En general, se observó que las terneras seropositivas al nacer incrementaron los niveles de anticuerpos después del calostrado (fig. 1a).
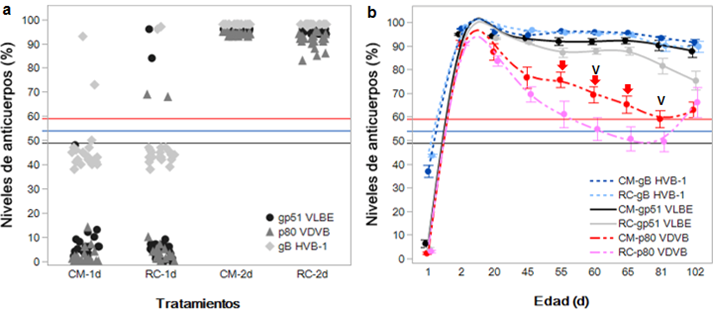
Figuras 1a y 1b: Niveles de anticuerpos frente a los virus de la leucosis bovina enzoótica, diarrea viral bovina y herpesvirus bovino 1. (1a) Transferencia pasiva de anticuerpos con base en el porcentaje de inhibición (PI) para la glicoproteína 51 del virus de la leucosis bovina enzoótica (VLBE), proteína no estructural NS3 o p80 del virus de la diarrea viral bovina (VDVB) y glicoproteína B de herpesvirus bovino 1 (HVB-1) (grupo CM=calostro materno, grupo RC=reemplazador de calostro). (1b) Cambios en los niveles de anticuerpos de las terneras alimentadas con CM o RC para el VLBE, VDVB y HVB-1. Las líneas horizontales marcan el punto de corte para animales seropositivos al VDVB (PI > 59, línea de color rojo), HVB-1 (PI > 54, línea de color azul) y VLBE (PI > 49, línea de color negro). Aplicación de CM intramuscular (tres dosis, flechas de color rojo) y vacunación (dos dosis, letra V de color negro). La barra de error indica el error estándar de la media.
Tabla 4: Medias de mínimos cuadrados de los niveles de anticuerpos frente a los virus en terneras alimentadas con calostro materno o reemplazador de calostro
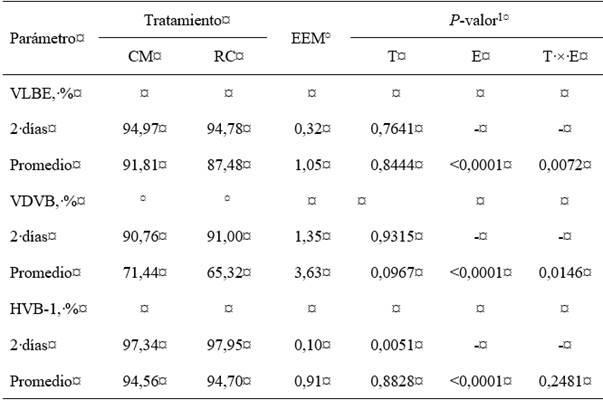
VLBE=virus de la leucosis bovina enzoótica; VDVB=virus de la diarrea viral bovina; HVB-1=herpesvirus bovino 1; CM=calostro materno; RC=reemplazador de calostro; EEM=error estándar de la media. 1T=efecto del tratamiento; E=efecto de la edad de evaluación; T × E=efecto de la interacción entre el tratamiento y la edad de evaluación
El análisis general de la media PI para el VLBE en cada evaluación mostró que todas las terneras conservaron un estatus seropositivo durante el ensayo, pero el grupo CM presentó una media superior en todas las evaluaciones después de los 45 días de edad en comparación con las medias PI del grupo RC, cuya diferencia fue más pronunciada a los 102 días de edad (fig. 1b); sin embargo, la comparación múltiple de las medias entre los grupos CM versus RC para un mismo punto de tiempo mostró que no hay diferencias significativas. Por otro lado, la diferencia entre las medias de los niveles de anticuerpos contra el VDVB y HVB-1 del grupo CM versus RC para una misma evaluación tampoco fueron significativas (tabla 4).
La aplicación de CM intramuscular no generó ningún cambio de tendencia en los niveles de anticuerpos frente al VLBE, VDVB y HVB-1 en las evaluaciones posteriores a los 55 días de edad ni tampoco la aplicación de la primera dosis de vacuna contra VDVB y HVB-1 (fig. 1b). También se apreció una diferencia en la media del PI para el VDVB desde los 20 hasta los 81 días de edad, periodo durante el cual las terneras dentro del grupo CM conservaron un estatus seropositivo, mientras las terneras dentro del grupo RC adquirieron un estatus seronegativo después de los 55 días de edad y seroconvirtieron después de los 81 días de edad como producto del estímulo antigénico generado por la vacunación (fig. 1b). Por su parte, las terneras con estatus seronegativo frente al VDVB aparecen primero en el grupo CM a partir de los 20 días de edad (fig. 2a).
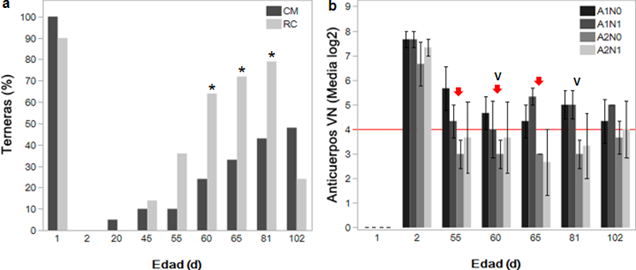
Figuras 2a y 2b: Inmunocompetencia frente al virus de la diarrea viral bovina en terneras. (2a) Proporción de terneras que alcanzan un estatus seronegativo frente al virus de la diarrea viral bovina (VDVB) cuando se alimentan con calostro materno (CM) versus reemplazador de calostro (RC) según el porcentaje de inhibición. (2b) Títulos de anticuerpos neutralizantes contra el VDVB según la prueba de virus neutralización durante el ensayo con CM intramuscular (tres aplicaciones, flechas en color rojo) y vacunación (letra V de color negro). La línea horizontal (color rojo) indica los títulos ≥ 1:16 o media log2 ≥ 4 asociados con la protección frente a la infección aguda. A1N0=grupo CM + sin aplicación de CM intramuscular; A1N1=grupo CM + aplicación de CM intramuscular; A2N0=grupo RC + sin aplicación de CM intramuscular; A2N1=grupo RC + aplicación de CM intramuscular. El asterisco indica diferencia significativa (P<0,05). La barra de error indica el error estándar de la media.
El análisis de regresión mostró que el porcentaje de terneras con estatus seronegativo se asoció mayormente con la ingesta de RC en comparación con el suministro de CM a los 60 (P=0,0159; 64,2 versus 23,8 %; respectivamente), 65 (P=0,0253; 71,4 versus 33,3 %; respectivamente) y 81 días de edad (P=0,0325; 78,5 versus 42,8 %; respectivamente) (fig. 2a). Por su parte, los títulos de anticuerpos medidos por la PVN y expresados como media log2 a los dos días de edad indicaron que la TP de anticuerpos neutralizantes distintos a los anti-p80 fue exitosa; sin embargo, a partir de los 55 días de edad, solo el grupo CM conservó niveles log2 > 4 asociados con la competencia inmune frente a la infección aguda, mientras las terneras dentro del grupo RC mostraron títulos menores (fig. 2b). Por otra parte, el efecto de la lactoterapia con CM intramuscular no generó cambios importantes sobre el PI frente a los virus o anticuerpos producto de la vacunación contra el VDVB debido a que la diferencia entre las medias log2 para los títulos de anticuerpos entre grupos A1N0 (4,5), A1N1 (4,5), A2N0 (3,2) y A2N1 (3,5) no fue significativa (P=0,1478 (fig. 2b).
Discusión
Todas las terneras incluidas en el estudio nacieron de vacas seropositivas al VLBE y HVB-1, lo que aumentó la probabilidad de registrar el nacimiento de terneras seropositivas o infectadas con alguno de estos virus según la literatura (Baillargeon et al., 2017; Mekata et al., 2021). Aunque HVB-1 no se detectó en circulación sanguínea, las evidencias demostraron el nacimiento de terneras seropositivas infectadas latentes (18,9 % (7/30)), terneras muy importantes como fuentes de infección para animales susceptibles durante el periodo neonatal (Petrini et al., 2019). Además, las pruebas precalostrales demostraron la ocurrencia de infección congénita y el nacimiento de animales seropositivos para el VDVB (8,1 % (3/37)), pero la proporción de animales recién nacidos con evidencias de la circulación del virus en sangre en hatos con infección endémica fue mayor (43,2 % (16/37)) a la proporción de terneras seropositivas (10,8 % (4/37)), contrariamente a lo planteado por Schefers et al. (2008).
En este estudio, la TP de anticuerpos contra el VLBE fue exitosa, pero Mekata et al. (2021) determinaron que la presencia de anticuerpos neutralizantes frente al VLBE solo evita la infección de forma parcial porque el número de animales infectados aumenta con la edad. Además, se observaron diferencias entre las medias para el PI asociado con el VLBE en diferentes puntos de tiempo cuando se comparó el grupo CM versus el grupo RC, es probable que la diferencia en la evolución de las medias se haya asociado con la carga proviral debido a que los títulos altos de anticuerpos indican alta carga proviral por la activación de la respuesta inmune para controlar la infección, en cambio las terneras dentro del grupo RC mostraron mayor resistencia natural para controlar la infección o mayor susceptibilidad que pudo haber favorecido el desarrollo de la enfermedad por los bajos niveles de anticuerpos (Forletti et al., 2020; Gutiérrez et al., 2014).
En este estudio, la TP de anticuerpos contra el VDVB y HVB-1 aumentó de forma significativa con ambos tratamientos; asimismo, Chamorro et al. (2014) reportaron tendencias similares para la transferencia de anticuerpos contra HVB-1 en terneros alimentados con CM o RC, pero la transferencia frente al VDVB fue mayor cuando los terneros se alimentaron con RC. Por su parte, la proporción de terneras seronegativas frente al VDVB aparecen primero en el grupo CM según el PI frente al VDVB, muy similar a lo descrito por Chamorro et al. (2014) . Sin embargo, la PVN indicó que las terneras dentro del grupo CM conservaron niveles de log2 > 4 y las terneras dentro del grupo RC presentaron títulos inferiores. Dado que un microtítulo sérico de 1:16 o un título medio de anticuerpos log2 de 4 por punto de tiempo se ha asociado con mayor susceptibilidad a la infección aguda causada por el VDVB (Chamorro et al., 2014), las terneras calostradas con RC presentaron mayor susceptibilidad para enfermar. En ese mismo sentido, la diferencia en el estatus sanitario con base en el PI de los anti-p80-DVB desde los 20 hasta los 81 días de edad y la PVN durante todo el ensayo indicaron diferencias en el desarrollo de la respuesta inmune humoral y celular, probablemente debidas a la diferencia en el potencial antigénico de las cepas contenidas en la vacuna versus las cepas de campo (Falkenberg et al., 2021) y al fundamento de las pruebas utilizadas (Hanon et al., 2017; Sangewar et al., 2020).
Además, el impacto de la infección aguda causada por el VDVB o HVB-1 no fue significativo, probablemente debido al efecto e interacción de factores asociados con la ubicación geográfica de la finca y con el programa de cría de las terneras; por ejemplo, la pasteurización del calostro, el suministro de CM de vacas multíparas, la alimentación con leche vendible y el alojamiento grupal en casos asociados con el CRB (Karle et al., 2019) y en los casos de diarrea, por su presentación multifactorial (Caffarena et al., 2021; Castells y Colina, 2021), en especial, por la asociación con otras etiologías distintas al VDVB, como rotavirus, coronavirus, Escherichia coli K99 y Cryptosporidium spp. (Brunauer et al., 2021) y al suministro de CM seropositivo frente al VDVB. Por otra parte, la lactoterapia con CM intramuscular no afectó la tendencia de los niveles de anticuerpos medidos por el PI frente a los virus o la concentración de anticuerpos neutralizantes como producto de la vacunación contra el VDVB, en contraposición al efecto favorecedor de la lactoterapia sobre la eficacia de las vacunas descrito por Prieto-Cristancho (2012).
Conclusiones
La TP de anticuerpos contra el VLBE, VDVB y HVB-1 fue exitosa en el grupo CM y RC, pero la duración de niveles asociados con la protección frente a la infección aguda causada por el VDVB fue adecuada solo para terneras alimentadas con CM. Sin embargo, el bajo porcentaje de diarrea y enfermedad respiratoria asociada con la presencia del VDVB y HVB-1, así como la recuperación de las terneras enfermas cuando el VDVB estuvo presente, sugirió que las terneras mostraron una inmunovigilancia adecuada para superar la infección debido a la TP adquirida con la ingesta de CM o RC, lo cual además evitó la reactivación de HVB-1 desde la latencia. Por lo que el uso de RC ante la ausencia de CM de alta calidad es viable. Por su parte, la aplicación de CM intramuscular no generó cambios de tendencia significativos sobre la respuesta inmune humoral frente a los virus.

