











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references

 Permalink
Permalink
Introduction
Infectious bovine keratoconjunctivitis (IBK) is a severe ocular disease that frequently affects livestock and has a significant economic impact worldwide (Cullen et al, 2017). The principal etiological agent of IBK is the Gram-negative bacterium Moraxella bovis (Henson y Grumbles, 1960). However, IBK outbreaks associated with the species Moraxella bovoculi have also been reported (Angelos et al, 2007). This disease is highly contagious and although it affects young animals more frequently, it is seen in bovines of different ages. Although IBK is rarely fatal, it generates significant economic losses associated with decreased vision, weight loss, diminished milk production, delays in growth, and high treatment costs and labor demand (Brown et al, 1998).
Several virulence factors have been associated with the pathogenicity of M. bovis, including fimbriae (pili), lipopolysaccharides (LPS), phospholipase B, outer membrane proteins (OMPs), acquisition of iron proteins, proteolytic enzymes as well as a hemolytic cytotoxin (Postma et al, 2008). It has been shown that M. bovis can express two different types of functional fimbriae, denominated type four pili I (TfpI) and type four pili Q (TfpQ), as a result of the inversion of a 2.1 kb DNA segment by site-specific recombination (Marrs et al, 1988). The capacity of M. bovis to produce more than one type of fimbria (phase variation) could increase antigenic variability and may result in antigenic switching, allowing M. bovis to evade the host immune response in vaccinated animals (Lepper et al, 1995; Postma et al, 2008). Several authors have also observed numerous serotypes of Moraxella spp. based on fimbrial antigens, which generate a marked antigenic diversity among strains. This diversity leads to challenges in the formulation of fimbriae-based vaccines (Moore and Lepper, 1991).
OMPs play a key role in microbial pathogenesis, virulence, and multidrug resistance, as well as in the mediation of many of the processes responsible for infection and disease progression (Jeeves y Knowles, 2015). In particular, OmpCD has been shown to act as a protective antigen in M. catarrhalis infections. OmpCD is conserved in most M. catarrhalis strains and exhibits attractive properties as a vaccine candidate (Hsiao et al, 1995; Saito et al, 2013). This protein is surface-exposed and contains at least two epitopes that are present in all of the studied strains of M. catarrhalis (Sarwar et al, 1992). OmpCD also exhibits characteristics of a porin-like OMP, relevant in nutrient acquisition (Holm et al, 2004; Saito et al, 2013). It has been proposed that M. catarrhalis OmpCD serves as an adhesin during human lung cell attachment acquisition (Holm et al, 2004) and plays an essential role in adherence to HEp-2 cells, contributing to the colonization and infection of laryngeal cells (Saito et al, 2013).
Mucosal vaccination has several advantages over traditional systemic vaccination, being the most effective route to elicit a protective response in mucosal surfaces (Holmgren y Czerkinsky, 2005). Given the mucosal location of the QIB, it is rational to consider the administration of an M. bovis vaccine by this route (Angelos et al, 2014).
The magnitude of the animal health problem caused by IBK has prompted researchers to generate effective vaccines for disease control. Currently, commercial vaccines are based on fimbriae or bacterins (inactivated whole bacteria) grown under conditions that optimize the expression of fimbriae (Brown et al, 1998). However, the protection elicited by commercial vaccines is often variable. The objective of this study was to produce a recombinant M. bovis OmpCD and evaluate its immunogenicity in a CD-1 mice model.
Materials and methods
Bacterial strains, plasmids, and growth conditions
Six Moraxella spp. strains comprising four Uruguayan field isolates (three M. bovis strains and one M. bovoculi strain) and two reference strains (ATCC Epp63 M. bovis and ATCC BAA1259 M. bovoculi) were used in the present study (Table 1). All isolates had previously been characterized in our laboratory (Sosa y Zunino, 2012). For the selection of the 6 strains, was considered the presence of 5 virulence genes that were previously detected in those strains: tolC, omp79, fur, mbxA and plB (Sosa et al, 2015). Moraxella spp. strains were routinely cultured in blood agar and incubated at 37°C. For the production of the recombinant protein, strains were routinely cultured aerobically in Luria-Bertani (LB) broth at 37°C; when solid media were required, 1.5% agar (LB agar) was added. When needed, ampicillin (100 µg/mL) was added to the media.
ompCD amplification by PCR and sequencing
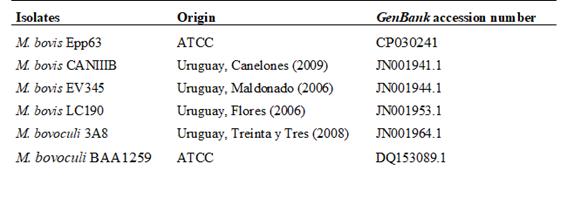
Genomic DNA of the different Moraxella spp. isolates was extracted using the Gen Elute Genomic DNA kit (Sigma) according to the manufacturer’s instructions. The following primers were designed according to the published sequence of M. bovis Epp63 genome (GenBank accession numberCP030241): OMPCD_IntF 5’-AAGCTCACGACAAACAACGTG-3’ and OMPCD_IntR 5’-TACGGTAGGTGCCAAACGAC-3’. Amplification was performed in a total volume of 25 μL, containing 1x PCR buffer, 1 U Taq polymerase (Invitrogen), 0.2 mM of each dNTP, 3.5 mM MgCl2, 0.5 μM of each primer, and 1 μL DNA (50 ng/mL). The reaction program included 30 cycles of 2 min to 94°C, 1 min to 59°C, and 1 min to 72°C. PCR products were sequenced in Macrogen (Korea). The sequences were edited using BioEdit and later using the BLAST software tool, and homology was evaluated using the NCBI public database. The nucleotide and amino acid sequences determined in this study were compared using MEGA software. Phylogeny trees were generated using the maximum likelihood method and the Kimura two-tailed model.
Expression and purification of M. bovis recombinant OmpCD
Genomic DNA of M. bovis EV345 was used to produce the recombinant protein. M. bovis EV345 is a strain that has been molecularly characterized in our laboratory. In addition to the presence of virulence genes, phenotypic characteristics such as twitching mobility are known associated to the pili type IV expression (Craig et al., 2004; Sosa et al., 2015). The primers were designed incorporating restriction sites sequences for NdeI and SacI (Invitrogen Anza) as follows:
OMPCD_NdeI_F:5´-CTTTGGAGGAACATATGAAACTTAATAAAATCGCTTTGG-3´, OMPCD_SacI_R:5´-GCCCTTGAGCTCTTAATGGTGATGGTGATGGTGTTTGGTTTCTACAACGTTAGTTAC-3´. The amplification reaction was performed using the same concentrations of the reagents described above. Cycling reactions were 30 cycles of 2 min to 94°C, 1 min to 63°C and 1 min to 72°C.
The pET system was developed for the cloning and expression of recombinant proteins in Escherichia coli. The target genes are cloned into the pET plasmids under the control of bacteriophage T7 transcription. In this work, the plasmid pET-21a (+) was used for the expression of the protein, which encodes for resistance to Ampicillin and presents the T7 promoter. After enzymatic digestion of the amplified EV345 ompCD and the pET21a(+) using the enzymes NdeI and SacI, products were purified and ligation was carried out using DNA ligase T4 (Invitrogen). The generated construct was first transformed into E. coli XL1-Blue and was then moved into the λ lysogen E. coli BL21(λDE3) by transformation, for expression of recombinant protein. Overexpression was performed under denaturing conditions according to the manufacturer’s instructions (Novagen, pET System) as previously described (Pellegrino et al, 2003). OmpCD expression was verified by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and Coomassie blue staining before and after induction of the T7 promoter with isopropyl-D-thiogalactopyranoside (IPTG). Prior to the purification of OmpCD, it was determined whether the protein was produced insoluble or insoluble form, according to the protocols defined by Novagen (pET System).
Protein purification was performed under denaturing conditions and then was eluted by nickel-agarose column (Pellegrino et al, 2003) assessed by 12% SDS-PAGE. Protein quantification was determined using Bradford’s method (Bradford, 1976). After electrophoresis, the OmpCD band was excised and sent to Institut Pasteur (Montevideo, Uruguay) for mass spectrometry analysis.
SDS-PAGE and Western blot
Recombinant OmpCD extraction and purification were monitored by 12% SDS-PAGE (Sambrook et al, 1989). Immunoblot analysis was performed using 1:200 tears and 1:100 sera from healthy and IBK-symptomatic calves. Tears and sera from a healthy animal and serum and tears from an animal with symptoms of IBK were tested. All samples were collected by veterinarians from VIRBAC Uruguay S.A. (Angelos et al, 2004). Proteins were transferred onto a nitrocellulose membrane (BioRad) for 1 h at 200 mA. Membranes were incubated overnight at 4°C in PBS 1x pH 7.4 with 5% skimmed milk. For antigen-antibody detection, membranes were incubated for 1 h at room temperature with a 1:30,000 dilution of goat anti-bovine IgG-alkaline phosphatase antibody (Sigma) (for sera) and 1:10,000 dilution of goat anti-bovine IgA-alkaline phosphatase antibody (Sigma) (for tears) (Sambrook et al, 1989).
Animals
Eight-week-old female CD-1 mice were used in the present study. Mice were purchased in “Division de Laboratories Veterinarios Miguel C. Rubino” (Dilave) and then they were kept in captivity conditions at the IIBCE breeding facilities. Mice were provided with food pellets and tap water ad libitum. All animal experiments were conducted in accordance with procedures authorized by the IIBCE Bioethics Committee (CEUA), Montevideo, Uruguay (protocol number 002/08/2015).
Subcutaneous immunization with recombinant OmpCD
A group of mice (n = 5) was subcutaneously immunized with 50 μg of the recombinant OmpCD combined with Freund’s complete adjuvant (Sigma) at a 1:1 ratio (300 μL total volume). Subsequent immunizations were performed under the same conditions but using incomplete Freund’s adjuvant (Sigma) 3, 7, 10, and 24 days after the first dose (Pellegrino et al, 2003). A second group (n= 5) of mice was treated with saline (300 μL) on the same days as negative control. Seven days after the last dose, the animals were anesthetized using ketamine (50 mg/kg) and xylazine (10 mg/kg) and were euthanized by cervical dislocation, the blood was extracted by cardiac puncture, and sera were obtained.
Nasal immunization with recombinant OmpCD
Four groups of mice (n = 6) each were intranasally treated with 50 µg of OmpCD, 50 µg of OmpCD combined with the mucosal adjuvant Carbigen (MVP Technologies, NE, USA), 50 µg of the mucosal adjuvant Carbigen, or saline. Vaccination was performed five times, on days 0 (first dose), 3, 7, 10, and 24. A volume of 30 μL was inoculated in each mouse (15 μL in each nostril) using a pipette fitted with a plastic tip, as previously described (Scavone et al, 2004). Blood was obtained from the caudal vein immediately before the first immunization (day 0). Seven days after the final dose (day 31), the animals were anesthetized using ketamine (50 mg/kg) and xylazine (10 mg/kg) and were euthanized by cervical dislocation. The blood was extracted by cardiac puncture, and bronchoalveolar lavage (BAL) was performed. The tracheae of the mice were exposed, and a small incision was made to introduce 1 mL of PBS and 1mM EDTA using a thin-tipped Pasteur pipette; three washes were performed to obtain the final product (Muñoz-Wolf et al, 2014). Blood samples were incubated for 1 h at 37°C and centrifuged at 3000 rpm for 15 min while BAL samples were centrifuged at 1000 rpm for 5 min. Sera and BAL supernatants were stored at -20°C until the experiments were performed.
Antibodies detection by enzyme-linked immunosorbent assay (ELISA)
OmpCD-specific IgG and IgA antibodies titers in serum and BAL fluid were determined by ELISA. Briefly, 96-well microtiter plates were coated with 50 µg/mL of recombinant OmpCD in 100 µL/well coating buffer (carbonate buffer, pH 9.4) and incubated overnight at 4°C. Plates were blocked with 200 µL/well of 20 mM Tris-HCl, pH 7.5, 200 Mm NaCl, 0,5 %, Tween 20, 3 % (p/v) skimmed milk, and incubated 90 min at 37 °C. Then, 100 µL of 1:4 BAL fluid and 1:100 serum dilutions were incubated for 90 min at 37°C. All samples were seeded in triplicate. Bound antibodies were detected by the addition of alkaline phosphatase-labeled goat antibodies specific for mouse IgG (whole molecule) or IgA ((-chain) (Sigma), diluted 1:30,000 and 1:40,000, respectively. Plates were incubated for 90 min at 37°C. After washing, p-nitrophenyl phosphate (Sigma) substrate (1 mg/mL) dissolved in diethanolamine buffer was added and plates were incubated for 45 min at 37°C. Finally, absorbance was measured at 405 nm using a Varioskan Flash multimode reader (Thermo Scientific).
Cytokines quantification
IFN-γ, IL-4, and IL-10 in the BAL fluid were evaluated by ELISA (OptEIA TM kit) containing the corresponding pair of specific monoclonal antibodies (capture and detection) against each cytokine according to the manufacturer’s instructions (BD Pharmigen, San Diego, CA, USA). A standard curve was generated based on serial dilutions of the cytokine recombinant standard proteins. Cytokine samples were interpolated in the standard curves. Absorbance was measured at 450 nm using a Varioskan Flash Multimode Reader (Thermo Scientific).
Statistical analysis
The basic assumptions of the parametric statistics were evaluated and the differences between the groups of animals were analyzed. In the cases in which the distributions met the assumptions, the analysis of variance (ANOVA) test was used to compare the means of the treatments. In the cases in which it was necessary to use non-parametric statistics, the comparison between groups was carried out using the Kruskal-Wallis test with the subsequent comparison between groups conducted using the Man-Whitney test. Statistical comparisons were carried out using the Past software with a confidence level of 95%.
Results
Detection of ompCD in different Moraxella spp. Strains
ompCD was detected by PCR in all the Moraxella spp. strains used in this study. The amplified products were sequenced, and their alignment revealed a great similarity between all the sequences, showing several internal conserved regions. The original published sequence of M. bovis Epp63 was included in the analysis. Regarding ompCD diversity among strains, allow nucleotide and amino acid variability between the M. bovis and M. bovoculi strains was observed (Fig. 1).
Expression of recombinant OmpCD in E. coli
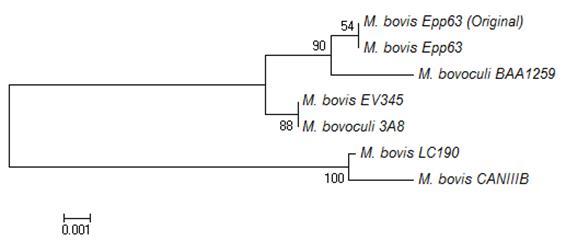
A notable band of approximately 45 KDa, corresponding to the predicted molecular weight of OmpCD (Fig. 2), was notice using SDS-PAGE in the total bacterial extract after induction, when assessed using SDS-PAGE. The OmpCD band was observed predominantly in the pellet obtained after centrifugation of the induced E. coli culture subjected to ultrasound treatment, corresponding to the insoluble fraction (Fig.2). The recombinant protein concentration was 3.33 mg/mL.
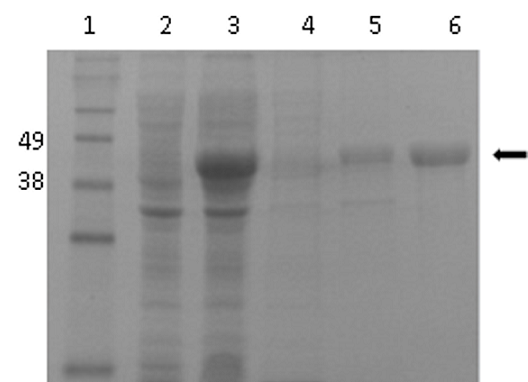
Figure 2: Recombinant OmpCD expression. Lane 1: molecular weight marker (KDa); lane 2: total bacterial protein extract before induction; lane 3: total bacterial protein extract after induction (expression of recombinant OmpCD); lane 4: soluble protein fraction after IPTG induction; lane 5: insoluble protein fraction after IPTG induction; lane 6: purified OmpCD eluted by a nickel-agarose column. The black arrow indicates the protein of expected size (45 KDa).
Identification of recombinant OmpCD
To corroborate the insert identity, the pET21a(+)-ompCD construct was extracted and the insert was sequenced. The insert showed an identity percentage of 99% with ompCD of M. bovis Epp63. After SDS-PAGE and Coomassie blue staining, the selected band was recovered for mass spectrometry analysis. This approach allowed the statistically significant identification of a hypothetical protein M. bovis. Likewise, this hypothetical protein is homologous to OmpA family protein M. bovis (NCBI Reference Sequence: WP_078273009.1) and OmpCD M. catarrhalis (GenBank: BAM17481.1).
Recombinant OmpCD immunogenicity
The Western blot showed the presence of OmpCD-specific antibodies in the sera and tears of diseased calves compared to healthy calves (Fig. 3), indicating that the recombinant OmpCD was recognized by antibodies of animals affected with IBK.
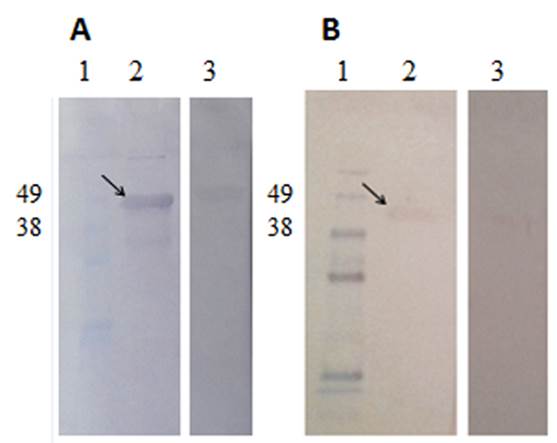
Figure 3: OmpCD reactivity in sera and tears of diseased and healthy calves by Western blot. Panel A: IgG response. Lane 1: molecular weight marker (KDa); lane 2: serum of a diseased animal; lane 3: serum of a healthy animal. Panel B:IgA response. Lane 1: molecular weight marker (KDa); lane 2: tears of a diseased animal; lane 3: tears of a healthy animal. The arrows indicate OmpCD, where the antigen-antibody reaction is visualized.
Antibody response in mice systemically immunized with recombinant OmpCD
The sera of mice systemically immunized with the recombinant OmpCD showed a significant induction of specific IgG, but not of IgA (Kruskal-Wallis, P<0.05) (Fig. 4).
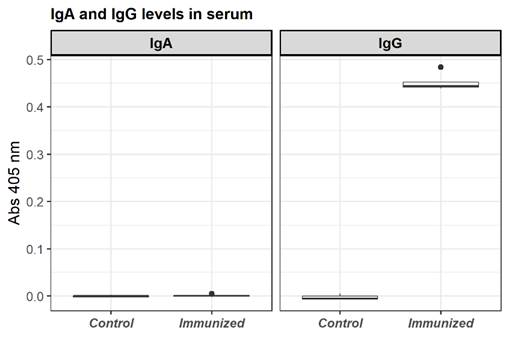
Figure 4: IgG and IgA levels in sera of mice immunized with OmpCD by the systemic route. In the left graph showed the IgA levels, Immunized: mice (n=5) treated with recombinant OmpCD; Control: mice (n=5) treated with saline. In the right graph showed the IgG levels, Immunized: mice (n=5) treated with recombinant OmpCD; Control: mice (n=5) treated with saline; e. The induction of IgG in immunized group compared to the control group is visualized. No induction of IgA is observed.
Antibody response in mice intranasally immunized with recombinant OmpCD
The induction of serum-specific IgG and IgA anti-OmpCD antibodies after intranasal immunization was not significant, either between the initial and final times within each group (Kruskal-Wallis, P>0.05) or among the different groups (Mann-Whitney, P>0.05).
The antibody response in BAL fluid was evaluated to compare IgG and IgA levels of the mice immunized by the intranasal route. IgG OD values were similar and generally low in the four groups of mice (Mann-Whitney, P>0.05). However, a significant induction of specific IgA anti-OmpCD was observed in the group immunized with OmpCD and the group immunized with the OmpCD combined with the adjuvant, compared to the groups treated with saline and with the adjuvant only (Mann-Whitney, P<0.05). There were no significant differences in IgA levels between the latter two groups (Mann-Whitney, P>0.05). The induced level of IgA in the group immunized with OmpCD combined with the mucosal adjuvant was significantly higher compared to the group immunized with OmpCD only (Mann-Whitney, P>0.05) (Fig. 5).
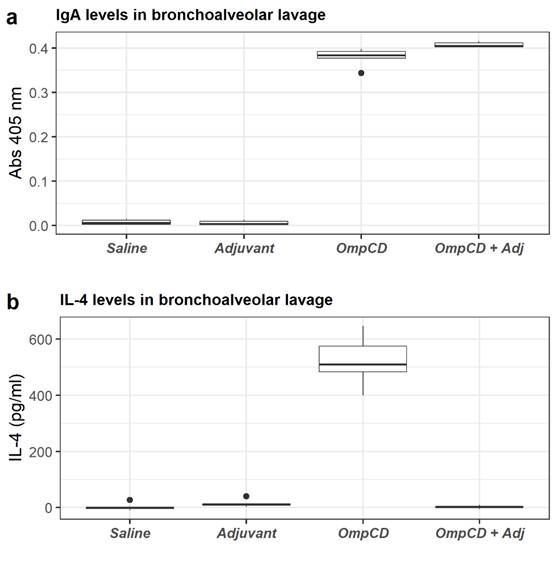
Figure 5: IgA and IL-4 levels in BAL fluid obtained from mice immunized with OmpCD by the intranasal route (Abs 405 nm). Panel a: IgA levels Saline: group 1, mice (n=6) treated with saline; OmpCD: group 2, mice treated with recombinant OmpCD; OmpCD+Adj: group 3, mice treated with recombinant OmpCD combined with the mucosal adjuvant; Adjuvant: group 4, mice treated with the mucosal adjuvant. Group 2 and 3 showed significantly higher values than groups 1 and 4. Panel b: IL-4 levels (pg/mL) Saline: group1, mice (n=6) treated with saline; OmpCD: group 2, mice treated with recombinant OmpCD; OmpCD+Adj: group 3, mice treated with recombinant OmpCD combined with the mucosal adjuvant; Adjuvant: group 4, mice treated with the mucosal adjuvant. Group 2 showed higher OD values than groups 1, 3 and 4
Cytokine response in BAL fluid of immunized animals
IL-4, IL-10, and IFN-γ levels were measured in BAL fluid obtained from all groups of mice immunized by the intranasal route using a commercial capture ELISA kit.
Significant induction of IL-4 was observed in the group of animals immunized with OmpCD compared to all of the other groups (Mann-Whitney, P<0.05) (Fig. 5). Surprisingly, mice immunized with OmpCD combined with the mucosal adjuvant did not show a significant response compared with the mice treated with OmpCD, with saline, or with the mucosal adjuvant only. IL-10 and IFN-γ levels were similarly low in the four groups (Mann-Whitney, P>0.05).
Discussion
It is generally desired that commercially available vaccines against Moraxella spp. prevent bacterial adhesion to the bovine corneal epithelium, a crucial stage in the infectious process. Therefore, efforts to develop an efficacious vaccine have primarily focused on the use of fimbriae to stimulate host immunity. However, the presence of multiple pilus serogroups (Moore y Lepper, 1991) together with the potential for pilin gene inversions (Marrs et al, 1988), increase antigenic variability and may result in antigenic switching. However, a high antigenic diversity of these surface organelles limits conferred protection (Lepper et al, 1995).
M. bovis possesses additional virulence determinants, including cell surface components such as OMPs. The most abundant bacterial OMPs are porins, which have a fundamental role in the host-pathogen interaction (Galdiero et al, 2012), which are highly conserved among isolates (Saito et al, 2013).
Interspecies recombination of M. bovis and M. bovoculi indicates that these species share a common gene pool. These observations suggest that there are potentially complex roles of both species in the etiology of IBK (Comin et al, 2020). Therefore, the design of a future IBK vaccine may benefit from the identification of conserved antigens shared by both Moraxella species, such as OMPs (Dickey et al, 2018).
Although OmpCD has been described as a potential antigen in vaccines in M. catarrhalis (Saito et al, 2013), it had not been characterized in M. bovis or in M. bovoculi to date. The use of vaccines against M. catarrhalis based on a native OmpCD antigens, unfortunately, not suitable for large-scale production undergood manufacturing practice conditions. Therefore, the production of OmpCD in a recombinant form is a valid alternative to produce this protein at a larger scale (Bertot et al, 2007).
Holm et al (2004) have proposed that a vaccine against M. catarrhalis containing OmpCD may interfere with adherence, a critical step in bacterial pathogenesis. Therefore, it could be suggested that immunization with OmpCD induce protective antibodies against other Moraxella spp. Systemic and mucosal immunization with recombinant OmpCD favored the lung clearance of M. catarrhalis in a mouse model (Murphy et al, 1998). Subsequently, Bertot et al (2007) observed that mice intranasally immunized with OmpCD combined with a mucosal adjuvant, developed an immune response that was able to promote efficient bacterial clearance after a pulmonary challenge with a virulent strain of M. catarrhalis, associated with a strong humoral and cellular systemic response (Bertot et al, 2007). Another study assessed the level of serum antibodies in children, observing that anti- M. catarrhalis OmpCD antibodies were inversely related to the severity of otitis media, suggesting a protective role in the outcome of the disease (Harabuchi et al, 1998).
We observed that ompCD gene was conserved within 4 clinical isolates of M. bovis and M. bovoculi from Uruguay and 2 ATCC reference strains. In contrast, the M. bovoculi pilin sequence is only 38% identical to the most similar pilin reported for M. bovis, suggesting that M. bovis pilin-based bacterins would provide limited protection against IBK (Calcutt et al, 2014).
Antibody reactivity against the recombinant M. bovis OmpCD was corroborated using sera and tears of diseased animals in Western blots. Intense bands were observed in diseased animals, while the presence of a lesser reaction in healthy animals could be explained since the animals were previously exposed to Gram-negative bacteria and in particular to Moraxella spp., which have been described as components of the normal ocular microbiota (O’Connor et al, 2012).
Pathogenesis and prevention of IBK have been investigated over many years, and the importance of local immunity to prevent IBK has frequently been highlighted (Angelos et al, 2014). Commercially available vaccines worldwide are systemically administered. Mucosal vaccines have the advantage of inducing immune responses at a general level but also locally (O’Sullivan y Montgomery, 2015). Therefore, the administration of an M. bovis vaccine could be performed using a mucosal route. The existence of a common and tightly interconnected mucosal-associated immune system enables immunization in one mucosal site to generate a significant immune response in a distant site (Haneberg et al, 1994). Different authors have suggested that vaccination by a route that stimulates mucosal IgA against M. bovis would be important to develop an effective vaccination strategy (Pugh et al, 1985).
In the present study, we used a murine model, as a low cost and useful research tool to preliminarily evaluate the ability of OmpCD to induce an immune response. We observed that intranasal immunization with recombinant OmpCD and recombinant OmpCD combined with a mucosal adjuvant (Carbigen) induced a significant specific IgA antibody response in BAL fluid. BAL fluid could be an appropriate driver to predict the induction of specific antibodies in tears, since it has been demonstrated that the ocular surface is an integral part of the mucosal immune system (Dua et al, 1995). Therefore, this approach was used to obtaining ocular secretions, which is not possible in mice. Conversely, it was observed that subcutaneous administration of the recombinant OmpCD protein produced only an increase in serum IgG.
Another strategy to improve the mucosal immune response is based on the use of adjuvants. In this work, the adjuvant Carbigen, which consists of an emulsion based on carbomer (Carbopol 934P), was used. Carbomer is a polymer that encapsulates the antigen, providing a slow release and depot effect in the presentation of the antigen to the effector cells, which significantly improves immune response and vaccine efficacy (Angelos et al, 2014; MVP Adjuvants, 2017). Addition of this adjuvant to the formulation showed a significant increase in antibody levels compared to the response induced by the protein only (without adjuvant). Therefore, the adjuvant improved the immunogenicity of the protein, and its incorporation in future vaccine formulations against IBK could be evaluated.
In the present study, the levels of INF-γ, IL-4, and IL-10 were evaluated in BAL fluid obtained from intranasally immunized mice. The predominant cytokine pattern determines the type of immune response induced (Palomo et al, 2002). Th1 lymphocytes can produce IL-2 and IFN-γ and mediate a cellular immune response. On the other hand, Th2 lymphocytes produce IL-4 and IL-10 among other proteins and mediate a humoral immune response (Abbas et al, 2008). The results obtained revealed that intranasal immunization with OmpCD induced a significant increase of IL-4 in BAL fluid. Cytokine concentration in BAL fluid of vaccinated mice indicated that cell-mediated immune mechanisms occurred in the host because of the exposure to M. bovis OmpCD. This result indicated that a Th2-type immune response was elicited after OmpCD immunization. Surprisingly, the group that received both OmpCD and adjuvant did not exhibit significantly higher IL-4 levels compared to the group treated with OmpCD, saline or adjuvant only. In the future, it would be interesting to evaluate a challenge in cattle and analyze the ocular secretions.
In conclusion, this is the first report about the possible role of M. bovis OmpCD as a
vaccine candidate against IBK. The results obtained in this work are encouraging and
provide the bases for the design of M. bovis challenge models in cattle.

