











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo

 Permalink
Permalink
Introducción
En la segunda mitad del siglo XX los criterios selectivos aplicados en la avicultura de carne se definieron en base al logro de una mayor producción, con máxima eficiencia en el uso del alimento y elevado rendimiento de los denominados cortes valiosos -pechuga y pata-muslo- lo que derivó en una óptima relación costo-beneficio y le otorgó a la actividad un perfil empresarial e industrial caracterizado por la integración vertical de las diferentes etapas del ciclo y el manejo intensivo de las aves en instalaciones de sofisticación creciente.
El mejoramiento genético llevado a cabo durante décadas, si bien redujo marcadamente el tiempo necesario para alcanzar el peso de faena mediante el aumento de la tasa de ganancia diaria de peso corporal, generó respuestas correlacionadas indeseables afectando el equilibrio que mantiene en armonía la etología y la producción animal, el ambiente y la diversidad genética, problemática que obligó a revisar progresivamente los criterios selectivos aplicados (Burt, 2002; Emmerson, 1997; Kerr et al., 2001; Pakdel et al., 2005).
Estas respuestas negativas que acompañaron a la selección por rápido crecimiento, conjuntamente con una creciente preocupación por cuestiones vinculadas con el bienestar, incrementaron el interés de los consumidores por sistemas alternativos de crianza asociados a un mayor compromiso con el ambiente y la salud de los animales que aseguraran la provisión de alimentos de calidad (Sundrum, 2001). Ante este nuevo contexto, se plantean diferentes modalidades productivas que tienen en común el empleo de poblaciones con menor tasa de ganancia diaria de peso, la cría al aire libre al menos durante una parte del ciclo y el objetivo de promover productos diferenciados respecto de los convencionales.
El pollo Campero INTA (Bonino y Canet, 1999) es un biotipo de ave para carne generado en Argentina, que representa una opción en tal sentido. El mismo, de crecimiento más lento que el ave industrial, se cría en semi-cautiverio y se faena a una edad más próxima a la madurez sexual. Si bien fue desarrollado para el autoconsumo de familias con condiciones básicas insatisfechas representa también una alternativa de diversificación para productores a pequeña escala, en aquellas situaciones en las cuales las opciones genéticas comerciales disponibles muestran desajustes ambientales, requiriéndose, entonces, un recurso genético capaz de adaptarse a esta situación.
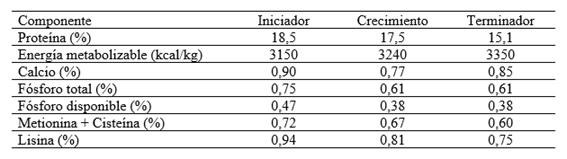
El esquema tradicional de alimentación de los pollos camperos incluye la utilización de tres tipos de raciones: Iniciador, Crecimiento y Terminador. Dado que la formulación de estos alimentos difiere de la utilizada en la avicultura industrial, a lo que se suma la dificultad para disponer en el mercado del alimento Crecimiento, los pequeños productores familiares enfrentan complicaciones en el manejo que podrían superarse utilizando un esquema de alimentación basado en la combinación de solo dos tipos de raciones: alimentos Iniciador y Terminador. Manejos similares han sido propuestos en la búsqueda de recursos de bajo costo, en poblaciones de aves criadas en semi-cautiverio en zonas tropicales de América Latina (Martínez-Pérez et al., 2017).
El objetivo de este trabajo fue caracterizar el comportamiento productivo y la calidad físico-química de la carne, de machos de dos cruzamientos experimentales de tres vías de pollo campero en comparación con Campero INTA tomado como genotipo de referencia, bajo dos manejos de la alimentación. En este sentido se puso a prueba la hipótesis que estas dos nuevas opciones de pollos son igualmente aptas para para ser utilizadas en sistemas avícolas alternativos y que el reemplazo de tres tipos de alimentos por sólo dos, combinados en cinco dietas diferentes, no modifica su desempeño productivo final.
Material y métodos
Aves
Se utilizaron machos pertenecientes a tres grupos genéticos: Campero Casilda (CC), Campero Pergamino (CP) y Campero INTA (CI), provenientes del cruzamiento entre gallos de la población sintética paterna AH’ y gallinas producto de los cruzamientos recíprocos entre las sintéticas maternas ES y A (CC: ♂ES x ♀A; CP: ♂A x ♀ES) y de la sintética materna E (CI: genotipo de referencia). Todas las poblaciones sintéticas mencionadas se generaron y mantienen en el Núcleo Genético de la Sección Avicultura en la EEA Pergamino de INTA. La constitución genética de dichas sintéticas es (Bonino, comunicación personal): Sintética A (75% Cornish Colorado 25% Rhode Island Red), Sintética ES (87,5% Cornish Colorado 12,5% Rhode Island Red), Sintética E (50% Cornish Colorado 50% Rhode Island Red) y Sintética AH’ (50% Hubbard 50% estirpe Anak grises).
Manejo
Las aves se produjeron en la Estación Experimental Agropecuaria (EEA) del Instituto Nacional de Tecnología Agropecuaria (INTA) en Pergamino, Argentina. Al nacimiento se sexaron por inspección de la cloaca, se vacunaron contra la enfermedad de Marek, se identificaron con bandas alares numeradas y se trasladaron (n = 120 por grupo genético) a las instalaciones de la Facultad de Ciencias Veterinarias de la Universidad Nacional de Rosario en la localidad de Casilda, provincia de Santa Fe, donde fueron sometidas al manejo establecido en el protocolo de producción de pollos camperos (Bonino, 1997), en un todo de acuerdo con las normas éticas exigidas por la Universidad Nacional de Rosario. Las aves se criaron inicialmente a galpón, como un único grupo, con una densidad inicial de 15 animales por m2 hasta los 28 días de edad. Durante ese lapso recibieron un fotoperíodo de 16 horas con un ciclo mínimo de oscuridad de 8 horas. Durante las primeras semanas de vida la temperatura ambiente del galpón se reguló a 26 °C (33ºC a la altura de los pollitos) disminuyendo paulatinamente a razón de tres grados por semana hasta llegar a 21 ºC. A partir del día 28, se seleccionaron al azar 60 aves de cada grupo genético las que se dividieron aleatoriamente en dos subgrupos de 30 individuos y se alojaron hasta su faena a los 84 días de edad, en seis galpones contiguos, con acceso a parque, con una densidad de 2 aves por m2.
Se implementaron dos manejos de la alimentación. El Grupo MT (n = 30 aves de cada genotipo) recibió el manejo tradicional para pollos camperos basado en la utilización de tres tipos de alimento (Iniciador, Crecimiento y Terminador). El Grupo MA (n = 30 aves de cada genotipo) fue sometido a un manejo alternativo basado en sólo dos tipos de alimento (Iniciador y Terminador). Los alimentos utilizados, formulados específicamente para pollos camperos de acuerdo al protocolo de producción, se ofrecieron ad libitum, al igual que el acceso al agua de bebida. La composición de los alimentos, formulados y preparados en la Sección Avicultura de la EEA Pergamino de INTA, se resume en la Tabla 1.
Las aves con manejo tradicional (MT) recibieron Iniciador entre el nacimiento y los 35 días de edad; Crecimiento entre los 36 y los 56 días de edad y Terminador entre los 57 días y los 83 días de edad. Las aves con manejo alternativo (MA) recibieron Iniciador (100%) entre el nacimiento y los 28 días de edad; mezclas de Iniciador (75%) + Terminador (25%) entre los 29 y los 42 días de edad; Iniciador (50%) + Terminador (50%) entre los 43 y los 56 días de edad e Iniciador (25%) + Terminador (75%) entre los 57 y los 70 días de edad y, por último, Terminador (100%) entre los 71 y los 83 días de edad. Durante el experimento todas las aves recibieron el plan sanitario indicado en el protocolo respectivo según el siguiente detalle de vacunaciones: día 1: Marek; día 14: Newcastle-Bronquitis infecciosa + Gumboro; día 35: Viruela; día 45: Newcastle-Bronquitis infecciosa + Gumboro (refuerzo) y el suministro de un antiparasitario de amplio espectro en el día 28.
Variables respuesta
En cada ave alojada a piso, considerada como una unidad experimental, se registró el peso corporal con aproximación al gramo a intervalos semanales entre el nacimiento y la faena. Los datos longitudinales peso corporal-edad cronológica correspondientes a 15 aves de cada subgrupo genotipo x manejo de la alimentación se ajustaron en forma individual con la función sigmoidea de Gompertz (Fitzhugh, 1976): Wt = A exp (- b exp (-kt)), donde: Wt = peso corporal (g) en el tiempo t; A = peso corporal maduro o asintótico (g); b = parámetro de posición, constante de integración sin valor biológico; k = tasa de aproximación al peso corporal asintótico A y t = edad en semanas. El ajuste se llevó a cabo por regresión no lineal utilizando una técnica iterativa (GraphPad Prism versión 2.0) basada en el algoritmo de Marquardt. La bondad del ajuste se evaluó en base a tres criterios: la convergencia de las iteraciones en una solución, del valor del coeficiente de determinación no lineal (R2) ajustado y la aleatoriedad en la distribución de los residuales (test de rachas). A los efectos de los análisis estadísticos los valores de los estimadores de peso corporal asintótico (A) y de tasa de maduración para peso corporal (k) correspondientes a cada ave se consideraron como nuevas variables aleatorias.
La uniformidad en peso corporal de cada grupo experimental mantenido a piso se estimó semanalmente durante todo el ciclo a partir del valor del coeficiente de variación (desvío estándar fenotípico relativizado por el valor del promedio del carácter y expresado como porcentaje (Toudic, 2007)). A partir de los 36 días de vida, noventa aves adicionales, treinta de cada grupo genético, a razón de 15 por modalidad de manejo, se alojaron en jaulas individuales (30 cm de frente x 42 cm de altura x 50 cm de profundidad) para el control del consumo voluntario de alimento. Luego de un período de acostumbramiento de siete días, cada ave se pesó semanalmente y se determinó su consumo diario individual de alimento entre los 42 y 84 días de edad. A partir de estos datos se calculó la eficiencia alimenticia (aumento de biomasa por unidad de alimento consumido) y la relación de conversión (cantidad de alimento requerida para aumentar un kg de peso) (Pym y Solvyns, 1979; Cahaner et al., 1987).
A los 84 días de edad, tras un ayuno de 12 horas, 20 aves elegidas al azar de su respectivo grupo genotipo x manejo de la alimentación criado a piso, se faenaron en la Sección Avicultura de la EEA INTA Pergamino, por corte neto de la vena yugular, previa insensibilización mediante dislocación cervical, con desangrado total en 1,5 minutos como mínimo. Se registró el peso vivo en ayunas, el peso eviscerado, el peso de la grasa abdominal (Griffith et al.,1978; Becker et al.,1984), el peso de la pechuga no deshuesada y el peso en conjunto de la pata y el muslo derechos. El rendimiento de la carcasa se calculó como el cociente entre el peso eviscerado y el peso vivo pre-faena. Se tomaron muestras de pechuga a cinco aves elegidas al azar de cada tratamiento (grupo genético x manejo de la alimentación), las que se enviaron adecuadamente acondicionadas en bolsa de polietileno de 1º uso a la Cátedra de Tecnología de los Alimentos de la Facultad de Ciencias Veterinarias de la Universidad Nacional del Nordeste, Corrientes, Argentina. Las muestras se mantuvieron congeladas a -18°C y a temperatura de refrigeración (2,5 ± 1ºC) una vez descongeladas 24 horas previas a las determinaciones. La calidad de la carne de ambos cortes procesados sin piel (análisis proximal: humedad, proteína bruta, extracto etéreo y cenizas), terneza, capacidad de retención de agua y pérdidas por cocción se evaluaron con los métodos analíticos convencionales establecidos por la American Meat Science Association (AMSA) (Cross, 1978) para el análisis físico y por la Association of Official Analytical Chemists (AOAC) (Latimer, 2012) para el análisis químico.
Análisis estadístico (Sheskin, 2011)
El efecto del grupo genético, del manejo de la alimentación y de la respectiva interacción simple entre ambos sobre las diferentes variables respuesta se evaluó con un análisis paramétrico de la variancia correspondiente a un diseño completamente aleatorizado con un experimento factorial 3 x 2 (tres grupos genéticos x dos modalidades de manejo) de acuerdo con el siguiente modelo:
Yijk = µ + αi + βj + (αβ)ij + eijk
dónde: Yijk = k-ésima observación correspondiente al ij-ésimo grupo
µ = efecto de la media general
αi = efecto fijo del i-ésimo grupo genético (i = 1-3)
βj = efecto fijo del j-ésimo manejo de la alimentación (j: 1-2)
(αβ)ij = efecto de la interacción grupo genético x manejo de la alimentación
eijk = error experimental
En todos los casos se consideraron estadísticamente significativas las comparaciones con probabilidad asociada menor al 5%.
En los casos en que interacciones significativas dificultaron la interpretación de los efectos principales el efecto del manejo de la alimentación, dentro de grupo genético, se evaluó con la prueba t de Student de comparación de medias para datos independientes y, el efecto del grupo genético para cada modalidad de manejo, con un análisis de la variancia a un criterio seguido de la prueba de comparaciones múltiples de Tukey.
Resultados
Los valores de los estimadores de los parámetros de la función de Gompertz con significado biológico correspondientes al ajuste de los datos peso corporal-edad cronológica en los diferentes tratamientos se presentan en la Tabla 2.
Tabla 2: Estimadores de los parámetros de la función de Gompertz en dos híbridos experimentales de tres vías de pollo campero y en el genotipo de referencia, bajo dos manejos de la alimentación.
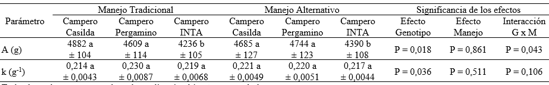
Todos los valores corresponden a la medida aritmética ± error estándar
Tamaño muestral: n= 15 aves por grupo genotipo-manejo de la alimentación
ab Valores con diferente letra difieren al menos al 5% para las comparaciones entre grupos genéticos dentro de manejo de la alimentación
A: peso corporal asintótico. k: tasa de maduración (velocidad de aproximación a A)
La Figura 1 muestra las trayectorias correspondientes al comportamiento del peso corporal de los diferentes grupos entre el nacimiento y la faena.
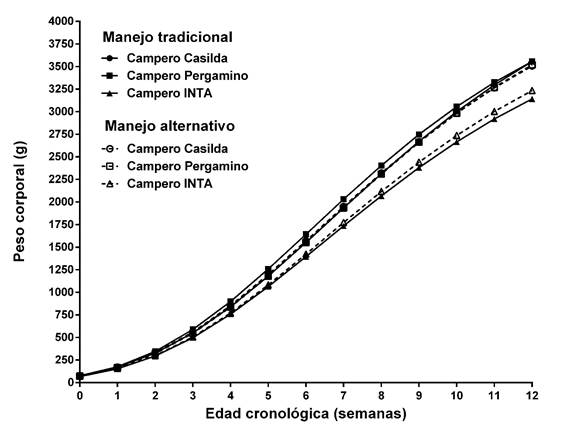
Figura 1: Curvas de crecimiento teóricas de dos híbridos experimentales de tres vías de pollo campero y del genotipo de referencia bajo dos manejos de la alimentación.
La ausencia de interacción grupo genético x manejo nutricional permitió evaluar los efectos de los factores principales. El tamaño asintótico de los grupos genéticos evaluados (CC y CP) fue similar entre sí y mayor al del genotipo de referencia (P < 0,001). Los genotipos no difirieron en la tasa de maduración. El manejo de la alimentación no afectó el valor de los estimadores de los parámetros. En CC el manejo alternativo tendió a disminuir el peso asintótico y a aumentar la tasa de maduración. En los dos grupos restantes el aumento de tamaño asintótico estuvo acompañado de una disminución de la tasa de maduración.
La Figura 2 presenta el comportamiento dinámico del coeficiente de variación como indicador de la uniformidad interna de cada grupo de aves.
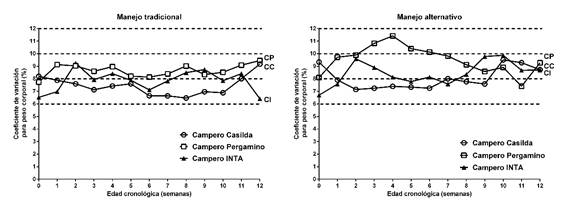
Figura 2: Uniformidad en peso corporal dos híbridos experimentales de tres vías de pollo campero y del genotipo de referencia bajo dos manejos de la alimentación.
Al evaluar el efecto del manejo de la alimentación sobre la uniformidad, dentro de genotipo, se observó una oscilación entre el 6% y el 10% en Campero Casilda y Campero INTA y entre el 8% y el 12 % en Campero Pergamino. Las aves con manejo alternativo de la alimentación tendieron a ser menos uniformes, hecho particularmente evidente en el caso de Campero Pergamino, genotipo en el que el grupo alimentado con dos dietas y sus combinaciones presentó valores mayores o iguales al 10% entre los 7 y los 49 días de edad.
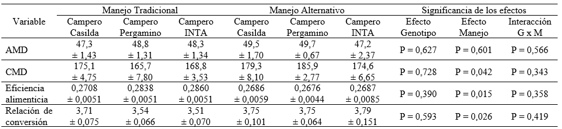
La Tabla 3 presenta los valores de aumento medio diario de peso corporal, consumo medio diario de alimento, eficiencia alimenticia y relación de conversión de alimento. No se observaron interacciones significativas.
Tabla 3: Relación de conversión y variables asociadas en dos híbridos experimentales de tres vías de pollo campero y en el genotipo de referencia, bajo dos manejos de la alimentación.
Todos los valores corresponden a la medida aritmética ± error estándar
Tamaño muestral: n = 15 aves por grupo genotipo-manejo de la alimentación.
P > 0,05 para todas las comparaciones entre grupos genéticos dentro de manejo de la alimentación
La eficiencia alimenticia y la relación de conversión al igual que los caracteres asociados (AMD y CMD) fueron similares entre genotipos. El manejo de la alimentación no afectó el AMD, pero el cambio del manejo tradicional a alternativo aumentó el CMD (P= 0,041) con disminución de la eficiencia alimenticia (P= 0,014) y, por ende, aumento de la relación de conversión (P= 0,030).
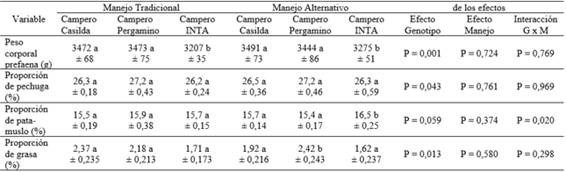
La Tabla 4 resume los valores correspondientes a los caracteres productivos evaluados a la faena.
Tabla 4: Caracteres productivos a la faena en dos híbridos experimentales de tres vías de pollo campero y en el genotipo de referencia, con dos manejos de la alimentación
Todos los valores corresponden a la medida aritmética ± error estándar.
Tamaño muestral: n = 20 aves por grupo genotipo-manejo de la alimentación
ab Valores con diferente letra difieren al menos al 5% para las comparaciones entre grupos genéticos dentro de manejo de la alimentación
El manejo de la alimentación no afecto a ninguna de las cuatro variables. El efecto del genotipo fue significativo en todas ellas (peso corporal: P < 0,001; proporción de pechuga: P= 0,043; proporción de pata muslo: P= 0,049 y proporción de grasa abdominal: P= 0,013). La interacción sobre la proporción de pata-muslo (P= 0,020) se debió a la mayor proporción del corte de CI bajo el esquema alternativo de alimentación. Las aves de ese grupo genético tendieron a ser algo más livianas a la edad de faena, diferencia que alcanzó significado en las comparaciones efectuadas bajo el manejo tradicional de la alimentación. CP tendió a presentar mayor proporción de pechuga tanto con el manejo tradicional como con el manejo alternativo por lo que, considerado en conjunto, el efecto del genotipo resultó significativo (P= 0,043). CI presentó menor proporción de grasa abdominal diferenciándose de ambos cruzamientos experimentales, en particular con el manejo alternativo de la alimentación. Bajo el manejo tradicional CC presentó mayor contenido de grasa que CP, situación que se invirtió con manejo alternativo, si bien, la variabilidad observada en el carácter explicaría la falta de significado estadístico de la interacción.
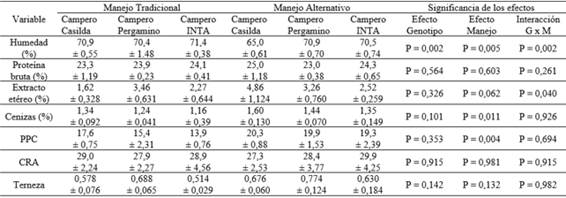
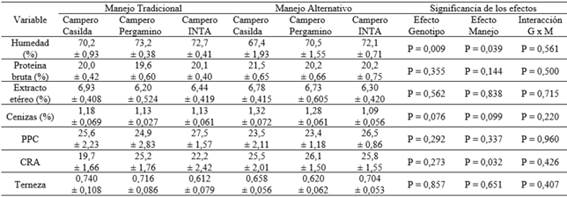
La Tabla 5 y la Tabla 6 presentan la composición química y los caracteres de calidad de la carne de pechuga y de pata-muslo, respectivamente.
Tabla 5: Composición química e indicadores de calidad de la carne de pechuga en dos híbridos experimentales de tres vías de pollo campero y en el genotipo de referencia, bajo dos manejos de la alimentación
Todos los valores corresponden a la medida aritmética ± error estándar.
Tamaño muestral: n = 5 aves por grupo genotipo-manejo de la alimentación
PPC: pérdidas por cocción - CRA: capacidad de retención de agua
Tabla 6: Composición química e indicadores de calidad de la carne de pata-muslo en dos híbridos experimentales de tres vías de pollo campero y en el genotipo de referencia, bajo dos manejos de la alimentación.
Todos los valores corresponden a la media aritmética ± error estándar.
Tamaño muestral: n = 5 aves por grupo genotipo-manejo de la alimentación.
P > 0,05 para todas las comparaciones entre grupos genéticos dentro de manejo de la alimentación a excepción del contenido de agua (menor en CC; P < 0,05)
PPC: pérdidas por cocción - CRA: capacidad de retención de agua
Con respecto a la composición química se observó n interacción (I) sobre el contenido de grasa (EE) de la pechuga (F=3,71; P=0,040) atribuible a un aumento del componente en las aves CC con manejo alternativo de la alimentación. En ninguno de los cortes se observaron efectos significativos del grupo genético (GG) ni del manejo de la alimentación (MA) sobre la magnitud del contenido de grasa y de proteínas. El manejo de la alimentación afectó el contenido de cenizas de las pechugas (P= 0 ,011) y la proporción de este componente fue levemente superior en ambos híbridos experimentales en el caso del corte pata-muslo (P= 0,096). En relación a los indicadores de calidad de la carne de pechuga, ninguno de los efectos analizados sobre la terneza mostró significado estadístico, si bien los híbridos experimentales de tres vías, en particular CP, mostraron una tendencia a presentar menor terneza de pechuga (mayor fuerza de corte) respecto del genotipo de referencia CI, con ambos manejos de la alimentación. Tampoco se observaron efectos significativos en la capacidad de retención de agua. En relación a las pérdidas por cocción, no se observó efecto de la interacción mientras que las pechugas de las aves sometidas a manejo alternativo de la alimentación presentaron, en promedio, valores más elevados (P= 0,004). En el caso del corte pata-muslo ninguno de los efectos analizados mostró significado estadístico ni en terneza ni en pérdida por cocción. En cuanto a la capacidad de retención de agua, no se observó efecto de la interacción ni del genotipo, pero, tal como se constató a nivel de la pechuga, se registraron valores más elevados (P= 0,032) en las aves sometidas a manejo alternativo de la alimentación.
Discusión
El análisis del patrón dinámico del peso corporal del pollo campero reviste importancia en tanto se lo destina a la producción de carne. La determinación genética del peso implica una base poligénica compleja (Bihan-Duval, 1997; Mignon-Grasteau, 1999), de naturaleza tanto aditiva, como lo evidencia la respuesta observada a la selección por este carácter, como no aditiva, con presencia de efectos heteróticos (Lariviere y Leroy, 2003) y maternos (Koerhuis y Thompson, 1997; Liu et al., 1993). La trascendencia de estos efectos difiere en los grupos evaluados en función de la estrategia genética que les dio origen. La respuesta productiva de CC y CP, por ser cruzamientos de tres vías y presentar como genotipo materno híbridos recíprocos entre dos poblaciones sintéticas, presenta componentes aditivos, de heterosis materna e individual como así también efectos maternos. CI es el resultado de un cruzamiento simple por lo que su desempeño productivo sólo incluye efectos aditivos y heteróticos individuales. Por su parte, desde un punto de vista ambiental, el manejo de la alimentación con un esquema simplificado como el propuesto, podría derivar en la presencia de interacciones genotipo x ambiente. La combinación particular de los valores de los estimadores de los dos parámetros -tamaño asintótico y tasa de maduración- que definen la forma de la curva de crecimiento, determinó que si bien, independientemente del manejo de la alimentación, los tres grupos genéticos crecieron con similar velocidad, CC y CP lo hicieron hacia un peso asintótico mayor, sin constatarse la asociación negativa esperada entre el tamaño asintótico y la tasa de maduración para peso corporal (Fitzhugh, 1976). Esta falta de asociación negativa, ya observada en estudios previos con aves camperas (Canet et al., 2011) pone en evidencia, al menos a nivel fenotípico, un comportamiento particular en la relación entre el peso maduro y la velocidad para alcanzarlo que podría atribuirse a cierto grado de independencia en los determinantes génicos de ambos caracteres. El esquema de alimentación alternativo, con sólo dos tipos de alimento combinados en cinco raciones, aun cuando afectó diferencialmente los valores de los estimadores de los parámetros en cada genotipo, no se tradujo en diferencias significativas en peso corporal entre las dos combinaciones experimentales, si bien determinó que CI crezca hacia un tamaño asintótico menor y presente menor peso corporal a la edad de faena.
Con respecto a la uniformidad en peso corporal, la disminución de la variación en el peso a la faena facilita el manejo en la planta de procesado y lotes con similares requerimientos nutricionales se traducen en menores costos de alimentación. Estudios anteriores en aves camperas (Lucach et al., 2013) mostraron que las mismas presentan, si bien con fluctuaciones a lo largo del ciclo de crianza, una buena uniformidad en los momentos críticos de dicho ciclo (nacimiento, traslado a parque y faena). Si bien estas poblaciones, criadas con menor control ambiental, presentan como característica una mayor variabilidad intrínseca, los resultados de este estudio evidencian que el manejo de la alimentación con dos raciones propuesto como alternativa, incidiría negativamente sobre el patrón de uniformidad de los lotes, hecho particularmente evidente en el caso de CP. Pese a ello, este comportamiento se revierte al finalizar el ciclo productivo, cuando todas las aves recibieron alimento Terminador, con similar uniformidad en el momento de la faena.
Se ha informado (Castellini, 2002, Wang, 2009) que la producción en sistemas extensivos resulta en un aumento del consumo de alimento con el concomitante deterioro de la relación de conversión en comparación con las aves criadas en sistemas convencionales. Los valores observados de relación de conversión, similares a los comunicados por otros autores en este tipo de aves (N’Dri et al., 2006) oscilaron entre un máximo de 3,79 y un mínimo de 3,51 para CI manejado con tres y dos raciones respectivamente. El manejo alternativo propuesto no modificó el aumento diario de peso de las aves, pero deterioró la relación de conversión al aumentar significativamente el consumo diario de alimento hecho posiblemente debido a que con acceso libre al alimento las aves requirieron mayor cantidad de una dieta no adecuadamente balanceada para expresar su potencialidad de crecimiento. Dadas las particularidades de estas poblaciones de crecimiento más lento que las empleadas en la avicultura industrial, con una edad de faena que prácticamente duplica a la del parrillero comercial, esta situación impacta negativamente sobre los costos de producción y justifica el mayor precio de venta de este producto diferenciado.
Los sistemas de producción alternativos a la modalidad de cría habitual de la avicultura industrial exponen a las aves a desafíos climáticos y a una mayor actividad física y, en consecuencia, se espera que maduren más lentamente retrasando la edad a la que alcanzan el peso objetivo de faena, particularidades que podrían traducirse en modificaciones en la calidad de su carne (Lawrie y Ledward, 2006). Si bien se ha comunicado escasa influencia del sistema de manejo en el rendimiento de carne a la faena (Blagojevic et al., 2009; Molee et al., 2011) existen opiniones contrarias en ese sentido (Issa Khan et al., 2017). Poblaciones de crecimiento lento en condiciones de producción orgánica mostraron una menor proporción de masa corporal, pero con superior calidad de carne, en relación a los sistemas convencionales de crianza (Grashorn y Serini, 2006). En contraposición, otros estudios con manejo en semi-cautiverio (Bogosavljević-Bošković et al., 2011; Wang, 2009) constataron mayor proporción de cortes valiosos -más relacionados con el peso corporal a la faena que con los factores nutricionales- y menor contenido de grasa abdominal, atribuido a la mayor movilidad de las aves que favorece la reducción del contenido de grasa corporal y estimula el desarrollo muscular. En relación a los valores promedio de proporción de pechuga y pata muslo observados en los grupos genéticos evaluados, los mismos fueron similares a los comunicados por Santos et al., (2005) a un peso objetivo de faena de 2500 g y superiores a los valores registrados por Farías et al., (2010) para machos de poblaciones comerciales en sistemas de crianza semi-intensiva, faenados a los 85 días. CI, tanto en el peso al sacrificio como en su peso maduro, tendió a ser más liviano que los genotipos experimentales. El manejo alternativo de alimentación no afectó la proporción de cortes valiosos, pero sí el contenido de grasa corporal, que fue menor en dicho genotipo y mostró un comportamiento inverso en los genotipos experimentales, ante esta modificación del ambiente nutricional, poniendo de manifiesto una interacción genotipo-ambiente.
Se ha jerarquizado la importancia del medio ambiente en que se encuentran las aves, el tipo de alimentación y la intensidad de actividad física, sobre la tasa de crecimiento, el tipo de fibra muscular que desarrollan y la deposición de grasa (Bancos, 2010; Bogosavljević-Bošković et al., 2010; Bogosavljević-Bošković et al., 2012; Erdal et al., 2012), lo que ejerce influencia en la composición y atributos sensoriales de la carne. El retraso en alcanzar el peso de faena, en comparación con el parrillero industrial conlleva a una mayor deposición de grasa lo que se traduce en un sabor más intenso de su carne (Fanatico et al., 2005), la que presenta mayor dureza por su mayor contenido de colágeno intramuscular, con mejores propiedades sensoriales, valores más elevados de pérdida por cocción y mayor jugosidad (Castellini et al., 2002). Se ha comunicado (Bogosavljević-Bošković et al., 2010, 2011) mayor contenido de proteína y menor proporción de grasa en cortes valiosos de aves criadas con acceso al exterior. Los resultados de este estudio evidenciaron que las aves con el manejo alternativo propuesto, presentaron mayores pérdidas por cocción en la pechuga, esperable en estos sistemas de crianza (Fanatico, 2006) y una mayor capacidad de retención de agua en el corte pata-muslo, parámetro habitualmente informado en bajos niveles en aves criadas en semi-libertad (Castellini et al., 2002). Independientemente del manejo, el contenido de proteína bruta observada en los cortes valiosos fue inferior al comunicado por algunos autores en aves con diferentes sistemas de crianza (Cheng et al., 2008) mientras que la proporción de grasa fue más elevada. No se observaron efectos significativos vinculados con el grupo genético ni con el manejo propuesto en el contenido proteico de los cortes valiosos, si bien CC tendió a presentar mayor proporción de proteína en las aves manejadas con manejo alternativo. La evidencia obtenida en este estudio confirma un mayor contenido de grasa (EE) y de cenizas en la pechuga de los cruzamientos experimentales en relación con el genotipo de referencia CI, resultados coincidentes con estudios previos en este tipo de poblaciones (Canet et al., 2013). El esquema de alimentación alternativo se tradujo en una mayor proporción de cenizas en ambos cortes, levemente superiores a los informados por otros autores para este tipo de sistemas productivos (Cheng et al., 2008; Bogosavljević-Bošković et al., 2010). Estas respuestas pueden resultar de interés desde el punto de vista nutricional y si bien el aumento consignado en extracto etéreo en pechuga resulta una desventaja en términos de eficiencia de conversión (Leclerq y Whitehead, 1988) al registrarse en el corte sin piel refiere al componente intramuscular, lo que podría traducirse en una mejor palatabilidad de un corte caracterizado por ser excesivamente magro.
Conclusiones
Los resultados indican que, si bien con particularidades en varios de los caracteres evaluados, no se observaron diferencias de trascendencia productiva entre los cruzamientos experimentales planteados como alternativa frente a Campero INTA. Campero Casilda y Campero Pergamino mostraron un desempeño equivalente en la mayoría de las variables analizadas y ventajoso en término de algunas de ellas -como es el caso del peso corporal- en comparación con el genotipo de referencia. Esta situación los posiciona como opciones para el tipo de producción avícola alternativa al que se los destina. El cambio progresivo propuesto en el manejo de la alimentación tampoco afectó, en términos generales, a los caracteres productivos evaluados, por lo que podría obviarse la inclusión de la categoría de alimento “Crecimiento” simplificando así su producción. El deterioro en la relación de conversión, de trascendencia en los sistemas intensivos, cobra menor importancia en este tipo de poblaciones caracterizadas por su menor eficiencia alimenticia y se ve compensado en este caso, por las ventajas derivadas de requerir sólo dos tipos de alimento.

