











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo

 Permalink
PermalinkIntroducción:
Las vacas de carne presentan una importante capacidad de convertir forrajes de baja calidad en proteínas de alta calidad para el consumo humano. No obstante, sólo el 5%del total de energía consumida durante el ciclo de vida es utilizado para la deposición de proteínas(Ritchie, 2000). Las principales razones de la ineficiencia de la producción de carne vacuna, se debería a la relativamente baja y lenta tasa de reproducción y el alto costo energético para su mantenimiento(Ferrell yJenkins, 1985).Los órganos vinculados al funcionamiento de los sistemas digestivo, respiratorio, circulatorio y actividades de excreciónconstituyen una menor proporción del peso del cuerpo (15-25%), pero no obstante representanmás del 50% de los costos de mantenimiento(Baldwin y col., 2004; Ferrell, 1988; Seal y Reynolds, 1993).Existe considerable evidencia que indica que la masa total de órganos difiere entre genotipos, entre estados fisiológicos y entre planos nutricionales (Baldwin, 1995; Baldwin y col., 2004;Casal y col., 2014; Ferrell, 1988;Jenkins y col., 1986) y que estas diferencias pueden traducirse en variaciones en las necesidades de energía de mantenimiento (Baldwin, 1995; Jenkins y col., 1986).
Considerando el uso global de la energía para los tejidos que la producción ganadera prioriza (ej. canal o musculo esquelético magro) se puede considerar a la energía utilizada por la masa total de los órganos como un “impuesto a la producción” (Reynods, 2002). El costo de mantenimiento tiene un impacto significativo en la partición de la energía metabolizable entre mantenimiento yproducción, por lo tanto puede afectar la eficiencia biológica y económica de la producción de carne (Johnson y col., 2003;Montaño-Bermudez y col., 1990).
El control de la intensidad de pastoreo a través del cambio en la oferta de forraje y el uso de genotipos cruza ha permitido mejorar la respuesta productiva y reproductiva de las vacas de cría (Carriquiry y col., 2012). Esta mejor respuesta productiva y reproductiva de las vacas se asocia a un mejor balance energético, esta mejora en el balance de energía podría ser el resultado no solo de un mayor consumo sino también de una reducción en los costos energéticos de mantenimiento (metabolismo basal y actividad). Estas respuestas diferenciales probablemente estén asociadas a diferencias en los mecanismos de partición de la energía (Carriquiry y col., 2012;Casal y col., 2014; 2017; Do Carmo y col., 2016;Soca y col., 2013).Es así que, una mejor comprensión de los factores asociados al costo de mantenimiento y balance energético de la vaca de cría pastoreando campo nativo mejoraría la comprensión de la eficiencia global del sistema de producción ganadero.
Este trabajo tiene como hipótesis que el control de la intensidad de pastoreo de campo natural, a través del manejo de la oferta de forraje, afecta la masa y composición de la canal así como de los órganos de vacas de cría puras (Hereford y Angus) y cruzas F1. Por lo tanto puede explicar - en parte - las diferencias en el comportamiento productivo-reproductivo entre diferentes ofertas de forraje y genotipos.El objetivo fue evaluar el efecto de dos ofertas de forraje (alta vs. baja) y el genotipo de los animales sobre el peso y composición de la canal y órganos en vacas de críapastoreando de campo natural.
Materiales y métodos:
El protocolo experimental fue aprobado y realizado de acuerdo con las normas de experimentación animal de la Comisión Honoraria de Experimentación Animal (CHEA) de la Universidad de la República.
Diseño experimental:
El experimento se llevó a cabo sobre 95 ha de campo natural (bioma Campos) en la Estación Experimental Bernardo Rosengurtt (Facultad de Agronomía, Universidad de la República, Uruguay, 32° S 54° W) desde junio 2007 hasta mayo 2010. Se utilizaron 32 vacas adultas (multíparas, 4 a 5 años) en un diseño de bloques completamente al azar con dos repeticiones en el espacio (según tipo de suelo: bloque 1: 60 ha suelo franco arenoso y bloque 2: 35 ha suelo franco arcilloso; cuatro parcelas en cada bloque) y un arreglo factorial 2 x 2 de oferta de forraje (OF) y genotipo de la vaca [GV, razas puras: Hereford (H) y Aberdeen Angus(A) y sus cruzas recíprocas F1 (HA y AH); PU vs. CR] .
La OF, se estimó como la relación entre la masa de forraje y la carga animal (kg de materia seca (MS) por kg de peso vivo (PV); Sollenbergery col., 2005) yrepresentó en el promedio anual 4 y 2,5 kg MS/kg PV(alta y baja oferta respectivamente, AOF vs. BOF) aunque fue variable a lo largo del año (5, 3, 4 y 4 kg MS/kgPV y 3, 3, 2 y 2 kg MS/kgPV para AOF y BOF en otoño, invierno, primavera y verano, respectivamente).La OF se ajustó mensualmente después de medir la disponibilidad de forraje en cada parcela (Haydock y Shaw, 1975) por el método de “put-and-take” (Mott, 1960). Las vacas experimentales se mantuvieron a lo largo del experimento en la misma parcela y en caso de ser necesario se añadieron o eliminaron vacas desimilar genotipo y estado fisiológico (“animales volantes”) basado en la disponibilidad de forraje.Los tratamientos de OF difirieron en masa de forraje y altura a lo largo del año(2072 vs. 1338 ± 160 kg MS/ha y 5.2 vs. 3.5 ± 0.3 cm de promedio anual para AOF vs. BOF, respectivamente), pero no difirieron (P ≥ 0.27) en su composición química[7.3, 8.6, 10.5 y 8.6 ( 0.5% PC y 42.6, 41.1, 36.2 y 40.2 ( 1.6 %FDA (base seca)en otoño, invierno, primavera y verano, respectivamente] .
Las vacas pertenecían a un grupo de animales experimentales generados como parte de un experimento dialélico de cruzamiento que se llevó a cabo durante 10 años en la Estación Experimental (Espasandiny col., 2010). Fueron evaluadas 8 vacas por tratamiento (AOF-CR, AOF-PU, BOF-CR, y BOF-PU; n = 4 para cada genotipo individual: H y A para PU o HA y AH para CR).Al comienzo del experimento (junio 2007) el PV y condición corporal (CC; escala de 1 a 8; Vizcarra y col., 1986) no difirieron(P > 0.20) entre los grupos (447 ± 58 kg y 4,1 ± 0.5 unidades, respectivamente). Las vacas pastorearon en la misma OF (AOF o BOF) y gestaron y lactaron un ternero cada año desde 2007 a 2010.
Muestreo y colecta de datos:
Al finaldel tercer añoexperimental (mayo 2010), a los 192± 10días postparto(45 días post destete) las vacasfueron sacrificadasen unmataderocomercial (PUL SA; CerroLargo,Uruguay, 40 km de laEstación Experimental). Previo al transporte, se registró el PV y la CC de todas las vacas.
Las vacas fueron aturdidas con una pistola de bala cautiva y luego fueron desangradas.El volumen de sangre se estimó de acuerdo a Hansardy col. (1953). Los animales desangrados se dividieron en cuero, extremidades, cabeza, tracto gastrointestinal (TGI), menudencias(tráquea, pulmones, corazón, diafragma, hígado, riñones), y canal. Los órganos de TGI y las menudencias fueron disecados y pesados individualmente. El peso de cada componente u órgano individual fue registrado y se recogieron muestras representativas que se almacenaron a -20°C para los análisis de composición química. Las muestras (100 a 200 g) fueron colectadas del TGI [retículo-rumen (~20 cm del esfínter pilórico, saco dorsal), omaso y abomaso (~20 cm del esfínter pilórico, curvatura menor), intestino delgado (~5 m unión ileocecal, yeyuno), intestino grueso (~2.5 m unión ileocecal, colon)] , vísceras rojas [pulmones (lóbulo apical), tráquea (~20 cm de la laringe),corazón (ventrículo izquierdo),diafragma (zona muscular y fibrosa),riñones (lóbulos caudales) ] ehígado (lóbulo dorsal). El peso de algunos órganos abdominales y pélvicos (bazo, páncreas, útero, vejiga) no se obtuvieron en su totalidad. El peso de la canal se registró antes y después del procedimiento de dressing. Secolectaron muestras de una sección representativa de la canal(media canal derecha) ubicada entre la 11ª y 13ª costilla (Hedrick, 1983). Esta sección se disecó en los tejidos blandos (tejido muscular, adiposo y conectivo) y el hueso, que se pesaron por separado y una muestra de los mismos se congeló a -20°C para la determinación de sucomposición química.
Análisis de composición química:
Las muestras de todos los órganos/tejidos se molieron en nitrógeno líquido y se analizó el contenido de agua (secadoa 105°Cen un horno deaire forzado durante48h), proteína (método Kjeldahl; proteína=nitrógenox6,25), lípidos totales (extracto etéreo;enextractor Soxhletdurante 20 h) y minerales (cenizas;incineración en horno mufla a 550°C durante16h) según normas de laAOAC (2000).La relación proteína:grasa se calculódividiendo la cantidad de proteína por la cantidad de grasa de los diferentes tejidos.
Cálculos y análisis estadísticos:
Luego del sacrificio, se estimó el peso corporal vacío (PCV) como el peso de la canalmásel peso total de extremidades, cabeza, cuero, sangre, órganos y vísceras (Hersomy col., 2004b). Los datos fueron analizados utilizando el paquete estadísticoSASSystemsProgram (SAS 9.0V; SAS Inst, Cary, NC, USA). Se utilizó el procedimiento UNIVARIATE para identificar valores atípicose inconsistenciasy para verificarla normalidadde los residuales.El análisis de los datos se realizó medianteel procedimiento MIXED,usando un modelo mixtoque considerabalaOF, el GVy su interaccióncomo efectos fijosy el bloquecomoun efecto aleatorio.La separaciónde medias se realizó mediantela prueba de Tukey. Se considerarondiferencias significativas, valores de P ≤ 0.05 y tendencias cuando0.05<P ≤ 0.10.Los resultados sepresentan comomedias de cuadrados mínimos±error estándar.
Resultados:
Peso vivo, peso corporal vacío ycondición corporal:
En promedio, durante los tres años de experimentación, el PV y CC fueron mayores (P ≤ 0.05) para vacas que pastorearon AOF vs. BOF y en las vacas CR vs. PU, siendo éstas diferencias más evidentes durante el último ciclo de gestación-lactancia (2009 - 2010, Figura I).
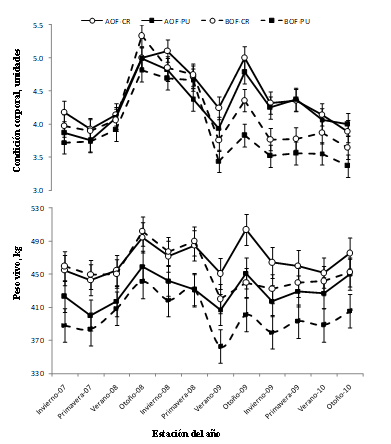
Figura I Peso vivo (A) y condición corporal (B) a lo largo de los tres años experimentales (2007-2010) de vacas puras (PU; Hereford y Aberdeen Angus; ■) y cruza recíprocas F1 (CR; ○),pastoreando alta (AOF; -) o baja (BOF; ---) oferta de forraje de campo natural (4 vs. 2.5 kg MS/kg PV, promedio anual). [n=32, 8 vacas por tratamiento] . Los datos se presentan como medias de cuadrados mínimos ± error estándar
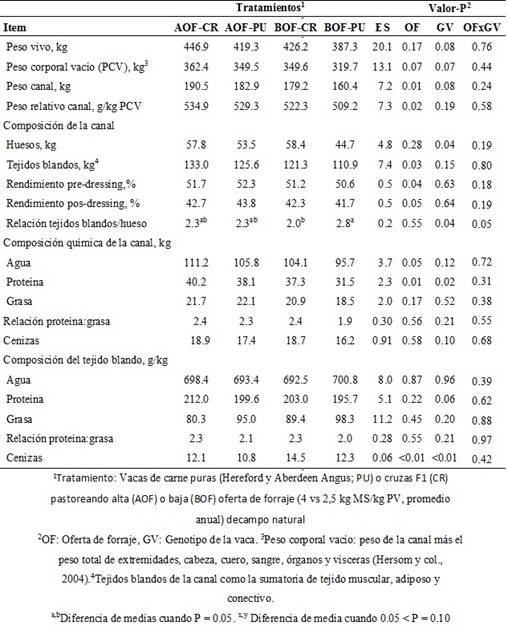
El PV de las vacas al momento de la faenatendió a ser mayor(P=0.08)en las vacas CRque en las vacasPU,mientras que el PCV tendió a ser mayor(P=0.07)en las vacas en AOF queen las de BOFy en las vacas CRque PU. Sin embargo, la CC de las vacas al momento del sacrificiono fue afectada (P ≥ 0.17) porla OF,el GVosu interacción(Cuadro I).
Canal y su composición:
El peso absoluto y relativo de la canal, los tejidos blandos (kg)y rendimiento pre y pos-dressing fueron mayores (P ≤ 0.05) en las vacas de AOF que en BOF (Cuadro I). El peso de la canaltendió a ser mayor (P = 0.08), presentado mayor (P = 0.04) proporción de hueso y menor (P = 0.04) relación tejidos blandos/hueso de la canal en las vacas CR que PU (Cuadro I). Sin embargo, la relación tejidos blandos/hueso de la canalfue afectada por la interacción entre la OF y el GV ya que las vacas BOF-PU presentaron un mayor relación que las vacas BOF-CR, siendo intermedia en las vacas CR y PU que pastorearon AOF (Cuadro I).
La cantidad de agua (kg) de la canal fue mayor (P = 0.05) en vacas que pastorearon AOF que en las que pastorearon BOF, pero la proporción (g/kg) de agua en los tejidos blandos no se vió afectada por la OF, el GV o su interacción (Cuadro I). La cantidad de proteína (kg) de la canal fue mayor (P ≤ 0.02) en vacas de AOF que en BOF y en vacas CR respecto a PU, pero en proporción (g/kg) en los tejidos blandos tendió (P = 0.06) a ser mayor en las vacas CR en comparación con las PU (Cuadro I). La cantidad de grasa en la canal o su proporción en los tejidos blandos, la relación proteína:grasa y la cantidad de ceniza en la canal no fueron afectados (P > 0.10) por la OF, el GV o su interacción (Cuadro I). La cantidad de cenizas (kg) en la canaltendió a ser mayor (P = 0.10) en las vacas CR que en las PU. Las vacas que pastorearon en BOF y CR presentaron mayor proporción (P ≤ 0.01) de cenizas en los tejidos blandos (g/kg) que las vacas que pastorearon en AOF y PU respectivamente (Cuadro I).
Peso absoluto y relativo a la canal de órganos y/o sección corporal:
El peso absoluto (kg) del cuero tendió a ser mayor (P = 0.08) en las vacas que pastorearon en BOF que en las que pastorearon en AOF. En contraste, el peso del TGI+hígado fue mayor (P = 0.02), mientras que los pesos del diafragma y los pulmones tendieron a ser mayores (P ≤ 0.09) en vacas que pastorearon en AOF que en BOF (Cuadro II).
Cuadro II Peso absoluto (kg) y relativo a la canal (g/kg canal) de órganos y secciones corporales de vacas de cría puras y cruza pastoreando distintas ofertas sobre campo natural
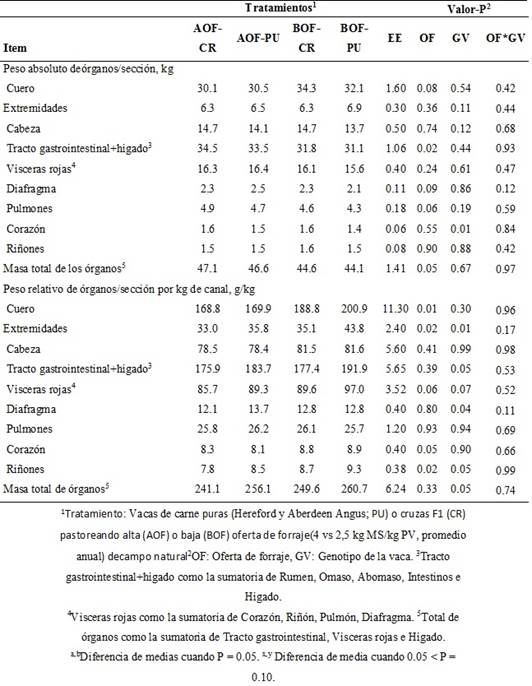
El peso de la masa total de órganos (kg) fue mayor (P = 0.05) en las vacas que pastorearon AOF que en las que pastorearon BOF. El peso absoluto del corazón fue mayor (P = 0.01) en vacas CR que en vacas PU. El peso absoluto de extremidades, cabeza, total de vísceras rojas y los riñones no se vieron afectados (P > 0.10) por la OF, el GV o su interacción (Cuadro II).
Los pesos relativos (g/kg respecto a la canal) del cuero, extremidades, corazón y riñones fueron mayores (P ≤ 0.05) en las vacas en BOF que en AOF. El peso relativo del total de las vísceras rojas tendió a ser mayor (P = 0.06) en las vacas en BOF que AOF, y tendió a ser mayor (P = 0.07) en vacas PU que en CR (Cuadro II). A su vez, el peso relativo de las extremidades, TGI+hígado, diafragma y riñones fue mayor (P ≤ 0.05) en las vacasPU que CR (Cuadro II). El peso relativo delamasa total de órganos fue mayor (P = 0.05) en vacas PU que CR. El peso relativo de la cabeza y los pulmonesno se vio afectado por la OF, el GVni su interacción (Cuadro II).
Composición química de los órganos:
El contenido de agua del cuero fue mayor (P = 0.01) en las vacas que pastorearon en BOF que en las que pastorearon en AOF, mientras que en los riñones tendió (P = 0.07) a ser mayor en vacas pastorearon en AOF que en las que pastorearon en BOF. El contenido de agua en la canal, diafragma, TGI+hígado, corazón y pulmones no fue afectada (P > 10) por la OF, el GV o su interacción (Cuadro III).
Cuadro III Composición química tisular de vacas de cría puras y cruza pastoreando distintas ofertas sobre campo natural
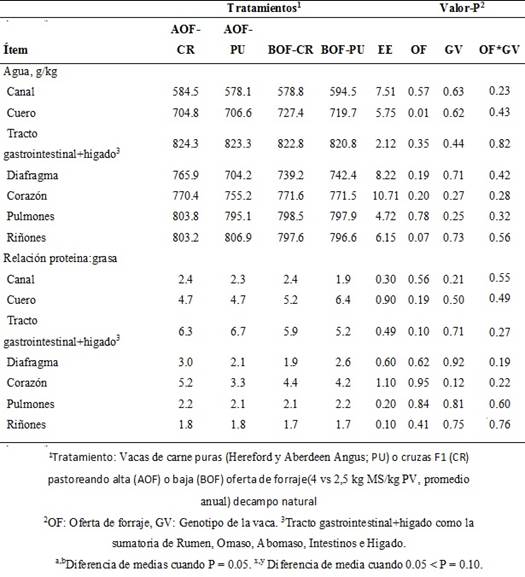
La relación proteína:grasa del TGI+hígado tendió (P = 0.10) a ser mayor en las vacas que pastorearon en AOF que en vacas que pastorearon en BOF. La relación proteína:grasa del resto de órganos, vísceras y la canal no fue afectada por la OF, el GV o su interacción (Cuadro III).
Discusión:
El presente trabajo identificó cambios en el peso y composición de la canal, así como en la masa de varios órganosdebido a cambios en la OF del campo natural o al GV que sugerirían una mayor eficiencia en el uso de la energía en vacas pastoreando en AOF que en BOF y para vacas CR respecto a PU.Estas diferencias podríantraducirse en variaciones en las necesidades de energía de mantenimiento, lo cual impactaríaen la partición de la energía metabolizable entre pérdidas de calor y energía neta de producción y por lo tanto, enla eficiencia biológica y económica de la producción de carne (Reynolds, 2002; Solis y col., 1988).
Efecto de la oferta de forraje:
Los menores PCVy peso absoluto y relativo de la canal de vacas que pastorearon en BOF se asociarían a una menor ingesta de energía (o menor ingesta de forraje).Do Carmo y col, (2016) estimaron el consumo de energía utilizando ecuaciones del NRC, donde reportaron un consumo de energía entre 10-12% más bajo en vacas que pastorearon en BOF que en AOF como consecuencia de una inferior masa de forraje.Dicha información resultó coincidente con lo reportado en bovinos para carne por Hersom y col. (2004a) donde los animales mejor alimentados presentaron un peso de canal mayor que los animales más restringidos. Además del mayor peso de la canal (+10%, AOF vs. BOF), los animales que pastorearon en AOF presentaron un mayor rendimiento de faena, mayor peso de tejidos blandos y mayor contenido proteico en la canal. Sin embargo, no se observaron diferencias en el contenido de grasa. Esto difiere con estudios previos (Houghton y col., 1990a; 1990b;NRC, 1996) donde una mejora en la alimentación y/o en nivel de energía en la dieta de vacas para carne incrementó el contenido de grasa corporal. En el presente trabajo, probablemente no se registraron diferencias en el contenido de grasa debido a que los animales pasaron a lo largo del experimento (3 años) por períodos de restricción y realimentación como se observa en la evolución de PV y CC(Figura I),y los mismos fueron sacrificados sin un periodo de terminación en un momento de baja acumulación de grasa (Casal y col., 2017).
El mayor PCV al sacrificio de las vacas en AOF se vio asociado, además de un mayor peso de la canal, a que las mismas presentaron o tendieron a presentar un mayor peso absoluto de la masa total de órganos. Investigaciones previas en novillos (McLeod y col., 2007; Wang y col., 2009) y vacas para carne (Meyer y col., 2012) reportan un aumento del tamaño del TGI con el aumento de la ingesta de alimento. En el mismo sentido, Jorge y col. (1997) observaron una reducción del peso de TGI e hígado en bovinos con restricción alimentaria. Las vísceras tienen altas tasas metabólicas (especialmente el hígado) y los órganos del TGI responden a cambios en la ingesta de alimentos. Kozloski y col. (2001) sugiere que el aumento de la ingesta conduce a un aumento de la actividad metabólica de las células epiteliales y de la actividad contráctil de las células musculares, determinando un aumento de la masa del TGI.
A pesar que el TGI e hígado constituyensolo el 10-13% del peso del cuerpo, estos tejidos representan más del 50% de los costos energéticos de mantenimiento, y por tanto contribuyende manera significativa a la variación en la productividad de los animales tanto dentro de un rodeo, asi como entre diferentes razas (DiConstanzo y col., 1991; Ferrell y Jenkins, 1985; Jenkins y col., 1991). Las vacas en AOF tendieron a presentar mayor peso de los pulmones y el diafragma, esto podría estar asociado a una mayor demanda de oxígeno a fin de metabolizar (oxidar) la mayor cantidad de nutrientes consumidos. Hentz y col. (2016) reportan en ovinos que a medida que se incrementa el nivel de ingesta de alimento, también aumenta el consumo de oxigeno por parte de la masa total de órganos.
En un estudio realizado por Jenkins y col. 1986 utilizando vacas de diferentes razas y con distintos niveles de alimentación concluyeron que los animales mejor alimentados y con mayor potencial de producción de leche presentaban mayor peso de los órganos del sistema respiratorio y circulatorio (corazón, pulmones y riñones) directamente relacionados a la mayor exigencia metabólica. En términos absolutos, la masa total de órganos acompasó con las variaciones de PV de los animales y fue mayor en vacas que pastorearon AOF. Estos resultados concuerdan con los resultados reportados por Burrin y col. 1992 donde expresan que la masa total de órganos es proporcional al nivel de consumo de alimento y al tamaño corporal de los animales.Contrariamente a lo que se esperaría, las vacas en BOF tendieron a presentar mayor peso del cuero, a pesar de que presentaban un menor tamaño corporal. Estas diferencias en el peso de este tejido estarían asociadas a un mayor contenido de agua en el mismo en vacas en BOF en comparación a las vacas en AOF.
Sin embargo, las vacas en AOF presentaron menor peso relativo a la canal del cuero y extremidadesque las que pastorearon en BOF. Ya que no se observaron diferencias en el peso absoluto, el menor peso relativo a la canal del cuero y extremidadesestaría relacionado con el mayor peso de canal de las vacas en AOF; una menor proporción de despojosrepresenta un aumento en el rendimiento de la canal. Vacas que pastorearon en BOF tendieron a presentar mayor peso relativo de las vísceras rojas (corazón y riñones principalmente), esto podría estar asociado a la mayor eliminación de urea y otros productos de desecho dado que éstaspresentarían una mayor movilización de reservas (grasa y proteínas; Casal y col., 2017). A su vez, los riñones (además del hígado) son tejidos querealizan neoglucogénesis, por lo tanto un aumento de la masa renal podría permitir una mayor suministro de glucosa (Reynolds y col., 2004). Contrariamente a nuestros resultados Wood y col. 2013 no encontraron diferencia en el peso de los órganos expresados como peso relativo a la canal entre vacas bajo distintos niveles de alimentación.Posiblemente,las diferencias en el tipo de dieta y al lago del experimento donde los animales pasaron por periodos de restricción y realimentación podrían estar explicando estas diferencias entre nuestro trabajo y lo reportado por Wood y col., 2013.
Efecto del genotipo de la vaca:
El PV, el PCV y el peso de la canal tendieron a ser mayores en vacas CR que en vacas PU, lo cual concuerda con lo reportado por Cundiff y col. 1974 y Morris y col. 1987, quienes atribuyeron la superioridad en PV de las vacas cruza a diferencias en el tamaño corporal o frame (tamaño del esqueleto). Esta diferencia en tamaño de la canala favor de las vacas CR se acompañó de una, mayor contenido proteico enla misma y en los tejidos blandos y presentaron mayor proporción de huesos. De manera similar, se han reportado efectos de la heterosis sobre el peso de la canal(Long, 1980). Gaines y col. (1967) evaluaron el uso de cruzamientos de razas británicas y reportaron valores de heterosis de 4% para el peso de la canal y para el área del músculo longissimus dorsi, pero no observaron diferencias significativas en el contenido de grasa de la canal. Contrariamente a nuestros resultados, Long y Gregory (1975) reportaron efecto de la heterosis no solamente sobre la cantidad de proteína (área del músculo longissimus dorsi) sino también sobre la cantidad de grasa en la canal.
Si bien las vacas CR presentaronun mayor peso de canal y contenido proteico en la misma, las vacas PU presentaronmayor relación tejidos blandos/huesos, específicamente las vacas BOF-PU presentaron una mayor proporción de dicha relación que las vacas BOF-CR.Estos resultados podrían serdebido a un mayor tamaño corporal, mayor frame, mayor estructura ósea y por lo tanto, mayor proporción de huesos en relación a la canalen las vacas CR. Esta mayor proporción de huesos se vio reflejado en una tendencia a un mayor contenido de cenizas (minerales) en la canal de las vacas CR. Wheeler y col. (1997) comparando diferentes cruzamientos reportaron valores superiores en el peso de los hueso en cruzas Salers (mayor frame) en comparación con cruzas Angus/Hereford (menor frame).
Los tejidos blandos de la canal de las vacas CR tendieron a presentar mayor contenido de proteínas y presentaron, al igual que las vacas en AOF, mayor contenido de cenizas en los mismos. Williams y col. (1983) reportaron mayor contenido de cenizas en los tejidos blandos de la canal y dentro de estas mayor contenido de zinc, fosforo, magnesio y potasio en estos tejidos en novillos terminados a forraje en comparación a los que fueron terminados con granos. Estos autores sugieren que este mayor contenido de minerales sumado a un mayor porcentaje de tejido magro (proteína) podría tener efectos beneficiosos en las características nutricionales de la carne.
Una mayor carga metabólica asociada a un mayorpotencial de producción de leche podría haber llevado a un incremento en las necesidades de transportar nutrientes y en la masa de órganos del sistema circulatorio, lo cual podría explicar el mayor peso absoluto de corazón en vacas CR que en vacas PU.En el presente estudio, la producción de leche durante la última lactancia (2009-2010; 140 d) fue mayor en las vacas de CR que en las de PU (Gutiérrez y col., 2013), asociadas con una mayor ingesta estimada de energía metabolizable(Do Carmo y col. 2016; Laporta y col., 2014).Solís y col. (1988) reportan mayores requerimientos de energía para mantenimiento en razas lecheras y sus cruzas vs razas carniceras, esto sería atribuido principalmente a un mayor tamaño de los órganos mas metabólicamente activos (TGI e hígado).
Ferrell y Jenkins (1985) informaron que las diferencias en las necesidades energéticas de mantenimiento entre los genotipos de vacas para carne se asociaron con el potencial de la producción de leche, que determinó las diferencias en la masa del hígado y de los órganos del TGI. Las vacas CR presentaron menor peso relativo a la canal del TGI+hígado lo cual estaría asociado a que estas tendieron a presentar un mayor peso de canal, una menor proporción TGIrepresentaría un aumento en el rendimiento de la canal. Además las vacas CR tendieron a presentar menor peso relativo de las vísceras rojas (espacialmente riñones), diafragma y particularmente de la masa total de órganos que las vacas PU. Particularmente, los costos demantenimiento varían entre vacas de diferentes genotipos. Estudios realizados por Solís y col. (1988) reportan que a pesar que animales Bos indicus y Bos taurus presenten un peso de canal y tasa de crecimiento similares, los animales Bos indicus y sus cruzas presentaron menores requerimientos energéticos para el mantenimiento, asociado a un menor tamaño del TGI, hígado y corazón y menor cantidad de grasa visceral. Varios trabajos coinciden en que el peso de los órganos difiere entre genotipos, entre estados fisiológicos y entre planos nutricionales (Baldwin, 1995; Jenkins y col., 1986;Solís y col., 1988;Smith y Baldwin, 1974) y que estas diferencias pueden traducirse en variaciones en las necesidades de energía de mantenimiento (Baldwin, 1995; Jenkins y col., 1986; Solis y col., 1988). Por lo tanto, nuestros resultados podrían sugerir que las vacas CR tendrían un menor costo de mantenimiento por unidad de producto que las vacas PU, dado fundamentalmente porque el peso relativo de la masa total de órganos respecto a la canal fue un 9.5% menor en vacas CR.Además, las vacas CR presentaron mayor proporción de grasa corporal, menor deposición de grasa visceral (omental/mesentérica) y mayor movilización de grasa y proteína en periodo de balance energético negativoque las vacas PU (Casal y col., 2014; Casal, 2017).
Conclusión:
El control de la intensidad de pastoreo de campo natural, a través del manejo de la oferta de forraje y genotipo de los animales afectaron la masa y composición de la canal y órganos. Las vacas en AOF y CR presentaron mayor peso de canal y mayor rendimiento carnicero y un menor peso relativo de los órganos respecto al peso de la canal. Estos resultados contribuyen a explicarlas mejoras en la eficiencia en el uso de la energía para producción, dado la menor relación entre masa total de órganos y canal.

