










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Links relacionados
Compartir

 Permalink
PermalinkVeterinaria (Montevideo)
versión impresa ISSN 0376-4362versión On-line ISSN 1688-4809
Veterinaria (Montev.) vol.51 no.197 Montevideo mayo 2015
Comparación de diferentes formulaciones de estradiol y de progesterona en un protocolo de sincronización de celos en vacas Holando en lactancia en un sistema pastoril; perfiles hormonales y respuesta ovárica
Comparison of different estradiol and progesterone formulations in an estrus synchronization protocol in lactating Holstein cows in a pasture-based system; hormonal profiles and ovarian response
Martínez-Barbitta Ma, González-Guasque Wb, Martínez-Piña Mb, Cavestany Dc*
a Doctor. en Ciencias Veterinarias (DCV), Programa de Posgrados, Facultad de Veterinaria, Universidad de la República, Montevideo, Uruguay
b DCV, Ejercicio liberal, Uruguay
c Doctor en Veterinaria (DV), MSc., PhD. Departamento de Reproducción, Facultad de Veterinaria, Universidad de la República, Montevideo, Uruguay
*Autor para correspondencia: Facultad de Veterinaria, Universidad de la República, Lasplaces 1620, 11700 Montevideo, Uruguay. E-mail: daniel.cavestany@gmail.com
Recibido: 6/5/2014 Aceptado: 28/7/2014
Resumen
El objetivo fue evaluar tres formulaciones de estradiol (E2) [5 mg de 17β (E17), 1 mg de benzoato (EB) y 1 mg de cipionato (ECP)] y dos presentaciones de la progesterona (P4) - dispositivo intravaginal (DIV) con 558 mg de P4 y 200 mg de P4 parenteral (MAD-4) - en un protocolo de sincronización de la ovulación. Se seleccionaron 30 vacas Holando en ordeñe que fueron presincronizadas con una combinación de GnRH y prostaglandina (PG) administradas con siete días de intervalo y que manifestaron celo en el mismo día; a los 10 días de éste se formaron 6 grupos (n=5 c/u): 1) DIV + EB: día 0: DIV + EB; Día 8: PG y retiro del DIV; Día 9: EB. 2) DIV + E17: día 0: DIV + E17; Día 8: PG y retiro del DIV; Día 9: E17. 3) DIV + ECP: día 0: DIV + ECP; Día 8: PG y retiro del DIV + ECP. 4) MAD-4 + EB: día 0: MAD-4 + EB; Día 8: PG; Día 9: EB. 5) MAD-4 + E17: día 0: MAD-4 + E17; Día 8: PG; Día 9: E17. 6) MAD-4 + ECP: día 0: MAD-4 + ECP; Día 8: PG + ECP. La respuesta fue evaluada por los niveles plasmáticos de E2, P4 y ecografía ovárica. Las concentraciones máximas de P4 se registraron una hora después del tratamiento (7,3±0,7 ng/mL DIV y 16,1±0,7 ng/mL MAD-4; P<0,0001); seis horas más tarde se redujeron a 5,7±0,7 ng/mL DIV y 11,2±0,7 ng/mL MAD-4 respectivamente (P<0.0001). Las concentraciones de E2 alcanzaron un máximo 4 horas después del tratamiento para grupos E17 y EB y 28 h después del tratamiento para grupos de ECP. Las concentraciones máximas fueron E17: 641,5±11,5 pg/mL, EB: 166,5±11,5 pg/mL y ECP: 45,6±10,9 pg/mL (P<0,0001). E2 volvió a los niveles iniciales 40 horas más tarde en grupos E17 y EB y 80 horas en grupos de ECP. Los diámetros foliculares en el día 8, fueron 14,9±1,4 y 14,4±1,3 mm para las vacas EB y E17 y 10,9±1,4 para los animales ECP (P<0,05). La ovulación ocurrió más temprano en los grupos EB y el porcentaje de vacas ovulando fue 88,9% en E17, 66,7% en EB y 30,0% en ECP (P<0,05).
Palabras clave: vacas lecheras, sincronización de celo, ovulación, estradiol, progesterona
Abstract
The objective was to evaluate three estradiol (E2) formulations [5 mg of 17β (E17), 1 mg of benzoate (EB) and 1 mg of cypionate (ECP)] and two presentations of progesterone (P4) - intravaginal device (DIV) with 558 mg of P4 200 mg of P4 parenteral (MAD-4) - in an ovulation synchronization protocol. Thirty lactating Holstein cows were presynchronized with a combination of GnRH and Prostaglandin (PG) given seven days apart and responding with a standing heat on the same day were selected; 10 days 6 groups were formed (n=5 each): 1) DIV+EB: Day 0: DIV + EB; Day 8: PG and DIV withdraw; Day 9: EB. 2) DIV+E17: Day 0: DIV + E17; Day 8: PG and DIV withdraw; Day 9: E17. 3) DIV+ECP: Day 0: DIV + ECP; Day 8: PG, DIV withdraw + ECP. 4) MAD-4+EB: Day 0: MAD-4 + EB; Day 8: PG; Day 9: EB. 5) MAD-4 + E17: Day 0: MAD-4 + E17; Day 8: PG; Day 9: E17. 6) MAD-4 + ECP: Day 0: MAD-4 + ECP; Day 8: PG + ECP. The response was evaluated by plasma levels of E2 and P4 and ovarian ultrasonography. The maximum average P4 concentrations were reached one hour after the treatment (7.3±0.7 ng/mL DIV and 16.1±0.7 ng/mL MAD-4; P<0.0001); six hours later were reduced to 11.2±0.7 ng/mL DIV and 5.7±0.7 ng/mL MAD-4 respectively (P<0.0001). E2 concentrations reached a maximum 4 hours after treatment for E17 and EB groups and 28 h after treatment for ECP groups. Maximum average concentrations were E17: 641.5±11.5 pg/mL, EB: 166.5±11.5 pg/mL and ECP: 45.6±10.9 pg/mL (P<0.0001). E2 returned to baseline levels 40 hours later in E17 and EB groups and 80 hours in ECP groups. The average follicular diameters at the Day 8 were 14.9±1.4 and 14.4±1.3 mm for the EB and E17 cows and 10.9±1.4 for the ECP animals (P<0.05). Ovulation occurred earlier in E17 and was 88.9% in EB, 66.7% in E17 and 30.0% to ECP (P<0.05).
Keywords: dairy cows, estrus synchronization, ovulation, estradiol, progesterone
Introducción
La disminución en la eficiencia reproductiva en vacas Holando de alta producción es un problema ampliamente reconocido y entre las razones se mencionan cambios hormonales que afectan la endocrinología reproductiva tales como reducción en niveles de insulina e IGF-I, que comprometen la función ovárica (Macmillan, 2010). En estos animales un aumento del metabolismo que incluye un incremento en la función hepática induce una degradación más rápida del estradiol que resulta en una reducción en la intensidad del celo (López y col. 2004, Wiltbank y col., 2006), lo que dificulta su detección. Es por eso que desde que se demostró que la ovulación se podía programar manipulando hormonalmente el eje hipófiso-gonadal (Pursley y col., 1995) una gran cantidad de protocolos que incluyen inseminación artificial a tiempo fijo (FTIA) han sido desarrollados en los cuales el control de la dinámica folicular y la ovulación han representado un avance fundamental en el desarrollo de tratamientos de sincronización de celos y de ovulación (Lucy y col., 2001).
Los primeros protocolos diseñados para sincronizar las ondas foliculares se basaban en el uso de GnRH (Pursley y col., 1995), el cual fue posteriormente sustituido por Estradiol (Colazo y col., 2003). Estos protocolos estimulan un celo natural cuando los estrógenos se aplican luego de la regresión luteal inducida por Prostaglandinas (PG) (Vynckier y col., 1990). Existe una amplia gama de preparados a base de estrógenos naturales o en sales sintéticas; éstas poseen absorción retardada desde el lugar de inyección y el estradiol 17β (E2) activo es liberado después de la hidrólisis. Cuánto más larga es la cadena del éster más baja es su solubilidad y más demora la absorción de la dosis completa. Por consiguiente, la vida media de las sales difiere acorde a su estructura (Souza y col., 2005); es así que el benzoato de estradiol (EB) tiene una vida media más corta que el cipionato de estradiol (ECP) (Bo y col., 2000).
En muchos protocolos de sincronización se emplea progesterona (P4) en forma de dispositivos intravaginales (DIV) con la finalidad de mantener exógenamente los niveles sanguíneos controlados y estables de esta hormona durante un tiempo determinados hasta su extracción (Colazo y col., 2002; Martínez y col., 2005) lo que ha resultado en una mejora en la fertilidad, tanto en vacas cíclicas como en anestro (Murugavel y col., 2003; Stevenson y col., 2006; Chebel y col., 2010). Sin embargo, el problema de la contaminación ambiental ocasionada por el deshecho de dispositivos de silicona con la mayor parte de la P4 luego de su utilización ha llevado al desarrollo de nuevas presentaciones de progesterona natural inyectable (Cavestany y col., 2008).
Se han evaluado varias sales de estradiol administradas al inicio de los tratamientos juntamente con dispositivos de progesterona, al día de la retirada de éstos o al día siguiente, con resultados variables (Burke y col., 2003; Souza y col., 2005). Sin embargo, no se han estudiado las correlaciones entre los niveles plasmáticos obtenidos con las diferentes drogas con el comportamiento reproductivo obtenido en los animales. Asimismo no se han presentado reportes que combinen diferentes sales de estradiol con diferentes formulaciones de progesterona. Por lo tanto, el objetivo del presente trabajo fue evaluar el efecto de la de dos presentaciones de progesterona y tres formulaciones de estradiol sobre los perfiles plasmáticos hormonales de E2 y P4, el desarrollo folicular y la ocurrencia de celos y ovulaciones en vacas lecheras presincronizadas con una combinación de GnRH y PG.
Materiales y métodos
Animales y tratamientos
Los procedimientos con animales fueron previamente aprobados por la Comisión de Ética en Uso de Animales (CEUA) de la Universidad de la República, Montevideo, Uruguay. El experimento se realizó en la Unidad de Lechería del Instituto Nacional de Investigación Agropecuaria (INIA) (La Estanzuela, Colonia, Uruguay). De 280 animales en ordeñe se seleccionaron 54 vacas multíparas, vacías y ciclando, en lactancia tardía (224±64 días), de condición corporal de 2,5±0,3 (escala de
Diez días antes del comienzo de los tratamientos las vacas se presincronizaron con GnRH (0,25 mg de Gonadorelina; Fertagyl, MSD Salud Animal, Sinervia, Montevideo, Uruguay) y, siete días más tarde, con 150 mcg de un análogo de PG (Prostaglandina, Laboratorio Río de Janeiro, Santa Fe, Argentina). Se colocaron parches para ayudar a la detección de celo (Estrotect, Gensur, Uruguay) y se realizó detección visual de celo durante media hora tres veces por día. Se seleccionaron 30 vacas que mostraron celo en un mismo día (48 horas luego de la administración de PG). El diseño experimental consistió en una combinación de las tres formulaciones de E2 y dos de P4. Para el tratamiento los animales se distribuyeron en 6 grupos equilibrados en base a DPP, CC y producción de leche (n=5 cada uno) y se utilizaron dispositivos intravaginales (DIV) de 558 mg de P4 natural (Cronipres M-24, Laboratorio Biogénesis-Bagó, Montevideo, Uruguay) o 200 mg de P4 natural inyectable (MAD-4, Laboratorio Río de Janeiro, Santa Fe, Argentina). Se utilizaron además tres formulaciones de estradiol: 1 mg de benzoato de estradiol (EB, Estradiol 10 Benzoato, Laboratorio Río de Janeiro, Santa Fe, Argentina), 5 mg de estradiol 17β (E17, Estradiol 17β Laboratorio Río de Janeiro, Santa Fe, Argentina) o 1 mg de cipionato de estradiol (ECP, ECP Estradiol, Laboratorios König, Montevideo, Uruguay) todos administrados vía intramuscular en el Día 0. La PG fue 150 μg de d-cloprostenol (PG; Prostaglandina, Laboratorio Río de Janeiro, Santa Fe, Argentina), por vía intramuscular. Los grupos experimentales quedaron formados de la siguiente manera:
- DIV+EB (n=5): Día 0: DIV + EB; Día 8: PG y retiro del DIV; Día 9: EB.
- DIV+E17 (n=5): Día 0: DIV + E17; Día 8: PG y retiro del DIV; Día 9: E17.
- DIV+ECP (n=5): Día 0: DIV + ECP; Día 8: PG + retiro del DIV + ECP.
- MAD-4+EB (n=5): Día 0: MAD-4 + EB; Día 8: PG; Día 9: EB.
- MAD-4 + E17 (n=5): Día 0: MAD-4 + E17; Día 8: PG; Día 9 E17.
- MAD-4 + ECP (n=5): Día 0: MAD-4 + ECP; Día 8: PG + ECP.
Se obtuvo sangre por punción yugular en tubos heparinizados al inicio del tratamiento de sincronización, hora 0 y a las horas 1, 6, 24, 48, 96, 144, 192, 196, 199 y 202 posteriores para determinar niveles plasmáticos de P4 (a la hora 192 se retiraron los DIV). Se realizaron sangrados adicionales cada 4 horas para determinar niveles plasmáticos de E2, desde el momento previo a la administración de los tratamientos hasta 68 horas para el E17 y EB y hasta 92 horas para ECP. La sangre fue centrifugada dentro de las dos horas de obtenida y el plasma separado y conservado a
Determinaciones hormonales
Las determinaciones hormonales se realizaron en el Laboratorio de Técnicas Nucleares de la Facultad de Veterinaria. La progesterona se analizó por RIA en fase sólida (125I Progesterona CAC, Siemens SA, Montevideo, Uruguay). La sensibilidad del ensayo fue de 0,02 ng/mL y los coeficientes de variación intraensayo para controles bajo (0,8 ng/mL), medio (8,0 ng/mL) y alto (15 ng/mL) fueron de 7,6%, 10,3% y 5,9%, respectivamente. La variación interensayo fue de 9,3%, 11,8% y 6,3% para los mismos controles. Para la determinación de estradiol se empleó RIA en fase líquida (125I Estradiol DA, Siemens SA, Montevideo, Uruguay) de acuerdo al método descripto por Meikle y col., (2001). La sensibilidad del ensayo fue de 1,4 pg/mL. Los coeficientes de variación interensayo fueron de 25,5%, 20,3% y 21,6% e intraensayo de 16,4%, 15,7% y 16,7% para controles bajos (7,8 pg/mL), medios (62,5 pg/mL) y altos (250 pg/mL), respectivamente.
Ultrasonografía
Desde el comienzo de los tratamientos de sincronización se realizó diariamente ultrasonografía ovárica por vía transrectal (Aloka SSD-500, Tokio, Japón) utilizando un transductor de 5 MHz para determinación del crecimiento folicular y presencia de cuerpo lúteo. A partir del D9 se realizó ecografía cada 4 horas para determinar el diámetro folicular máximo y el momento de la ovulación.
Detección de celo
Desde el D7 y durante 15 minutos cada 4 horas se realizó detección visual de celo. Asimismo se volvieron a colocar dispositivos de ayuda de detección de celos (Estrotect, Gensur SRL, Uruguay).
Análisis estadístico
Para el análisis de variables continuas con más de una medición durante el transcurso del experimento (concentraciones hormonales y crecimiento folicular) se utilizó un modelo mixto: PROC MIXED del SAS (Littell y col., 1998), que incluyó como efectos fijos: tratamiento, día e interacciones. La estructura de covarianza utilizada fue AR (1) y las medidas repetidas se realizaron sobre la unidad vaca anidada dentro de tratamiento por día. La ocurrencia de la ovulación se analizó por el PROC GENMOD de SAS (SAS Institute Inc., Cary NC, USA). El área bajo la curva, como medida global de los perfiles de liberación plasmática de las formulaciones evaluadas, se calculó por el método de trapezoides (Matthews y col., 1990). Se determinó el nivel de significancia en 5%.
Resultados
Estradiol
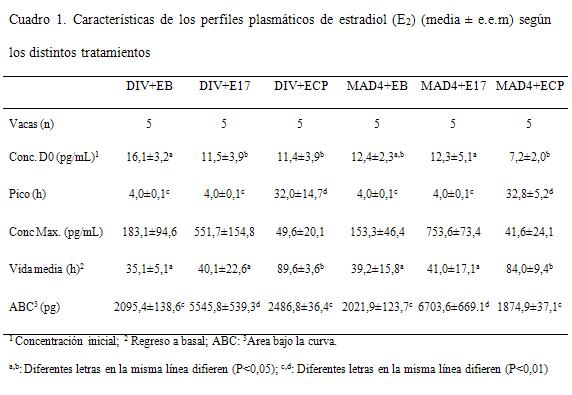
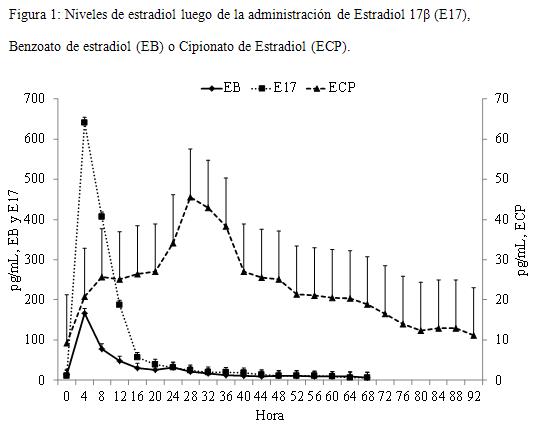
La concentración de E2 a la segunda dosis (D9 en E17 y EB, D8 en ECP) fue en promedio de 11,6±6,6, similar para todos los grupos (P>0,1). El pico de E2 se detectó a las 4 horas luego del tratamiento para los grupos E17 y EB y a las 32 horas para los grupos ECP (P<0,0001). Los valores máximos en la concentración también fueron distintos para las tres formulaciones de E2 ya que se registraron valores promedio de 652,6±66,1 pg/mL para los grupos E17, 169,8±72,3 pg/mL para EB y para los grupos ECP de 45,6±22,6 pg/mL (P<0,001). Luego del pico las concentraciones en los grupos E17 regresaron a los valores iniciales a las 16 horas manteniéndose así hasta el final del período. En los grupos ECP el pico fue menos pronunciado y las concentraciones descendieron 8 horas después (Cuadro 1 y Figura 1).
Progesterona
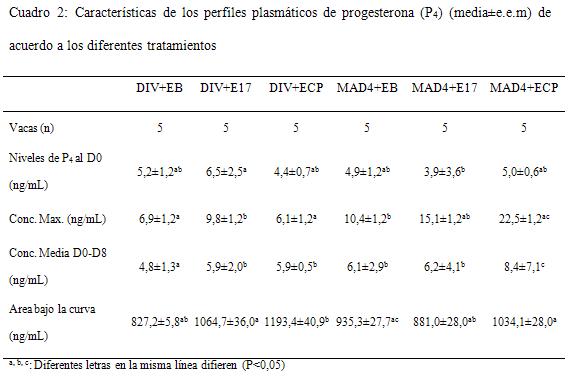
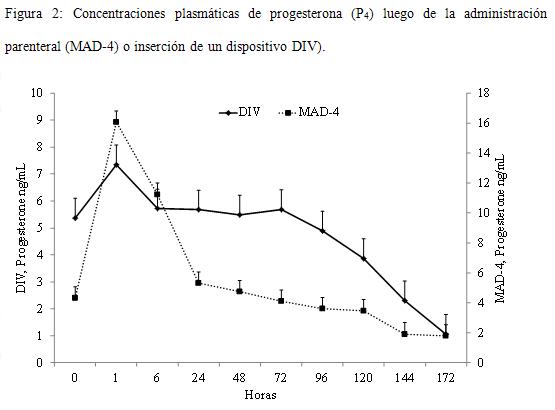
Las características de los perfiles plasmáticos de P4 entre los días 0 y 8 se presentan en el Cuadro 2 y la Figura 2. Los niveles plasmáticos al comienzo del tratamiento fueron de 4,4±0,3 y 5,2±0,3 ng/mL para DIV y MAD-4 respectivamente (P>0,1). Las concentraciones máximas se detectaron una hora después del tratamiento en los grupos con progesterona inyectable y seis horas después en los grupos con DIV. El valor de P4 al pico fue de 7,3±0,7 para DIV y 16,1±0,7 para MAD-4, (P<0,0001) y seis horas más tarde se redujeron a 5,7±0,7 y 11,2±0,7 ng/mL para DIV y MAD-4 respectivamente (P<0,0001), posteriormente manteniéndose en niveles similares hasta D8. Los niveles plasmáticos de P4 descendieron hasta niveles basales más rápidamente en las vacas tratadas con MAD-4 que con DIV, con un promedio entre grupos de 164,4±56,4 h para MAD-4 y 178,3±46,4 h para DIV.
Desarrollo folicular, ovulación y manifestación estral
Las características de crecimiento folicular, celo y ovulación se presentan en el Cuadro 3. El diámetro folicular al inicio de los tratamientos de P4 fue de 14,9±1,4 mm y 14,4±1,3 mm para EB y E17, y de 10,9±1,4 para los animales ECP (P<0,05). El diámetro folicular al momento del tratamiento con PG (D8) fue en promedio de 10,4±0,9 mm sin diferencias entre grupos y para el D9 (tratamientos E17 y EB) fue de 12,8±1,5 mm, 11,0±1,4 mm y 10,7 para EB, E17 y ECP respectivamente (P>0,1).
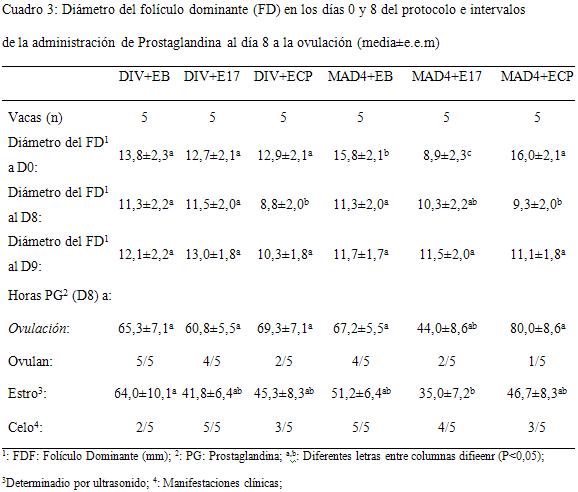
La luteólisis se determinó por la caída de P4 a los niveles previos a la administración o colocación de los dispositivos y ocurrió en el 77,8% de las vacas del grupo EB (7/9), en el 33,3% de las del grupo E17 (3/9) y en el 20% de las del ECP (2/10) (P > 0,1).
La ovulación (desaparición del folículo dominante a la ecografía) se detectó en el 9/10 de las vacas EB, 6/10 de las E17 y 3/10 de las ECP (P<0,05) y se produjo a las 60 horas después del tratamiento con PG, siendo de 63±3,7 h, 52±4,2 h y 66,6±6,0 h para las vacas en los grupos EB, E17 y ECP respectivamente (P>0,05). El tamaño del folículo dominante al momento de la PG fue significativamente más grande (P<0,05) en aquellos animales que ovularon, 10,8±2,3 mm vs. 8,8±2,5 en los animales en los que no desapareció el FD.
El comportamiento estral se manifestó en el 70% de las vacas del grupo EB, el 100% de las del E17 y el 60% de las vacas del ECP (P>0,1).
Discusión
La adición de E2 en tratamientos de inducción de ovulación produce un aumento rápido de la circulación de E17 con lo que se inhibe de la liberación prematura de PG y por lo tanto la expectativa del mantenimiento normal en la vida del cuerpo lúteo en protocolos IATF (Mann y Lamming, 2000). En protocolos Ovsynch modificados la adición de 0,5 o 1 mg de E17 8 h antes de la GnRH produce un aumento de esteroides circulante similar al natural, superior a las vacas control (Vynckier y col., 1990; Sartori y col., 2002; Wolfenson y col., 2004). Con estos supra niveles de concentración de E17 se puede esperar que se produzca mayor motilidad uterina y del oviducto (Hawk 1975), aumento del flujo sanguíneo uterino (Krzymowski y col., 2004) y aumento de la inmunidad local por mayor fagocitosis (Frank y col., 1983). Estos cambios podrían mejorar el ambiente uterino y podrían resultar en efectos positivos sobre la fertilidad.
La respuesta a las diferentes formulaciones de estradiol varió de acuerdo a las mismas. En las vacas tratadas con EB y E17 se registró un pico plasmático de E2 a las 4 horas del tratamiento, mientras que en las tratadas con ECP el mismo se observó a las 32 horas. El pico de E2 en respuesta al tratamiento con E17 fue cuatro veces más grande que con EB, mientras que el pico luego de ECP fue sensiblemente menor. Esta disímil liberación plasmática dinámica se aprecia claramente en el área bajo la curva (ABC) donde asimismo se constata la relación entre dosis administrada y niveles globales encontrados (la dosis de E17 es 5 veces superior a EB y ECP), donde la correcta elección en la frecuencia y posición de los sangrados, permitió diferenciar claramente los distintos perfiles de liberación plasmática del estradiol. Las diferencias en los perfiles sanguíneos de estradiol luego de tratamientos con diferentes estrógenos son consistentes con resultados previos (Vynckier y col., 1990; Bo y col., 2000; Haughian y col., 2002; Burke y col., 2003; Martínez y col., 2005). Souza y col. (2005) reportan valores más bajos pero utilizaron animales de más del doble de producción de leche que en este trabajo. Estos autores también reportan diferencias en los niveles de esteroides según la presencia o no de folículos dominantes, eliminando su producción natural de estradiol. La diferencia con el presente trabajo podría deberse a la influencia de la secreción endógena, ya que se utilizaron vacas ciclando que generaría mayores picos en las concentraciones de esta hormona así como mantenimiento de los niveles por más tiempo. Muy diferente fue el comportamiento de ECP donde el pico fue de 45 pg/mL a las 32 horas, llegando a niveles basales recién entre las
Los animales del presente ensayo fueron seleccionados tomando el criterio de inclusión según ciclicidad reproductiva lo que se constató hormonalmente con posterioridad y fueron presincronizados de modo que al empezar el protocolo se encontraran en fase luteal, con lo que los niveles de P4 fueron superiores a 3 ng/mL. Luego del tratamiento se registró un aumento en los niveles plasmáticos de P4 el cual fue mayor luego de la administración de MAD-4, que además presentó el pico de concentración antes que los dispositivos intravaginales. A partir de dicho momento los niveles de P4 cayeron más drásticamente en vacas tratadas con MAD-4 que con DIV; considerando que ambos productos contienen P4 natural, es muy posible que la vía parenteral no logre mantener niveles plasmáticos elevados por más de 24 h aunque esto también debería ser evaluado tratando animales en anestro o en fase folicular, de manera de evitar interferencia con P4 endógena de CL en vacas ciclando. Rogan y col. (2007) diseñaron un experimento con el fin de caracterizar perfiles de liberación de P4 plasmática de distintos dispositivos intravaginales con cantidades variables de hormona. Emplearon vacas lactantes ciclando, presincronizadas con PG. Encontraron gran equivalencia entre los picos de concentración (ng/mL) entre grupos (Cue-Mate: 5,0±1,1; DIB: 3,9±0,6; CIDR-
Los porcentajes de vacas que registraron luteolisis son coincidentes con aquellos encontrados por Callejas y col. (2008) con EB donde con 2 mg encontró 73,7% de regresión del CL y con 5 mg 85,6% y expresan la necesidad de inclusión de PG en protocolos con esteroides. Más dramáticas fueron los hallazgos en los casos de E17 y ECP no superando en ninguno de estos casos porcentajes de regresión del CL del 35%.
Se ha descrito la importancia de una concentración mínima circulante de E2 para la causar expresión del estro (Lammoglia y col., 1998). Lyimo y col. (2000) reportan una alta correlación entre los niveles séricos de E2 y la conducta estral en vacas Holando en lactancia. La adición de esteroides en protocolos de sincronización colabora en la expresividad conductual y es claramente diferencial acorde a los perfiles de liberación de las formulaciones. En nuestro ensayo encontramos gran diferencia en las demostraciones conductuales con un 100% de animales con expresión de para E17, mientras que esta cifra desciende a 70% y 60% en EB y ECP, respectivamente, que coincide asimismo con los niveles máximos plasmáticos obtenidos y también se corresponden con el tiempo para alcanzar la máxima concentración sanguínea, así en promedio las vacas E17 ya a las 4 h alcanzan su mayor concentración sanguínea de E2 con valores cercanos a 650 pg/mL y en su totalidad demuestran celo, mientras que las vacas EB y ECP logran el cenit plasmático a las 32 h con valores promedio de 170 y 45 pg/mL respectivamente que se relativiza en su menor expresión estral. En todos los casos los niveles de P4 al momento de la segunda dosis de E2 se hallaba por debajo de 1 ng/mL, luego de la remoción de los dispositivos a las 192 h (en los grupos correspondientes) y la aplicación de PG, siendo valorados promedio y no disimiles entre grupos de 1,4±0,7 ng/mL entre la remoción y la hora 202. Lammoglia y col. (1998) demostraron que las dosis que resultaron en celo inducido transitoriamente superaron los rangos fisiológicos lo que se relaciona directamente con los niveles plasmáticos, tal cual se vio en los resultados de este ensayo. Souza y col. (2005) mostraron que con dosis de 0,5 mg (EB, ECP y E17) el comportamiento estral no se manifestaba, excepto con E17, mientras que con 1 mg la respuesta conductual con montas e hiperactividad se presentaba entre las 12 y 36 horas en una proporción alta de los animales tratados, particularmente en aquellos tratados con E17 y EB. En la ocurrencia de la ovulación también se encontraron diferencias importantes entre los esteroides evaluados con diferencias significativas entre el EB con 89% de ovulación, frente a 67% y 30% para E17 y ECP respectivamente. Existe una relación de tendencia entre las horas al pico de concentración plasmática y los niveles encontrados con el momento de la ovulación, siendo anterior en E17, seguido de EB y finalmente ECP.
Cutaia y col. (2003) trabajando en vacas de carne con EB encontraron porcentajes de ovulación similares aplicando ECP tanto al momento de retirar el DIV como 24 h después y por los antecedentes encontrados con esta sal se procedió a administrar al mismo momento que la PG en vez de hacerlo al día siguiente como se efectuó con EB y E17.
El primer tratamiento con ECP podría no ser efectivo en sincronizar de la nueva onda folicular ya que al momento del segundo tratamiento con ECP la maduración del folículo sería variable o no completa y por lo tanto con escasa la respuesta ovulatoria (Bo y col, 2000). Las concentraciones plasmáticas por mayor tiempo en los animales ECP podrían haber suprimido la liberación endógena de FSH por intervalos más prolongados (Colazo y col., 2003) y de forma más variable en comparación con E17 y EB que conduciría a una aparición tardía de la onda folicular al inicio del tratamiento y menor tasa ovulatoria a la inducción con estradiol.
Conclusiones
Los resultados obtenidos sugieren que, en las condiciones de este ensayo, el E17 sería el compuesto que genera mejor respuesta conductual al estro, mientras que el EB sería ligeramente más efectivo en promover la ovulación y ambos mejores que el ECP. Hay que tener presente que éstos resultados pueden tener relación con las formulaciones de E2 administradas al inicio del protocolo. En cuanto a la P4, los dispositivos intravaginales generan respuestas más moderadas, estables y prolongadas que la P4 inyectable. Más investigación con estos productos en diferentes condiciones es necesaria para determinar estos efectos.
Referencias
1. Bo GA, Adams GP, Pierson RA, Tribulo HE, Caccia M, Mapletoft RJ. (1994). Follicular wave dynamics after estradiol-17β treatment of heifers with or without a progestogen implant. Theriogenology 41:1555-1569.
2. Bo GA, Bergfelt DR, Brogliatti GM, Pierson PA, Adams GP, Mapletoft RJ. (2000). Local versus systemic effects of exogenous estradiol-17 beta on ovarian follicular dynamics in heifers with progestogen implants. Anim Reprod Sci 31:141-157
3. Burke CR, Mussard ML, Gasser CL, Grum DE, Day ML. (2003). Estradiol benzoate delays new follicular wave emergence in a dose-dependent manner after ablation of the dominant ovarian follicle in cattle. Theriogenology 60:647-658.
4. Callejas S, Echevarria S, Zapiola A. (2008). Uso de benzoato de estradiol y un dispositivo intravaginal con progesterona en vacas cola de parición con servicio natural. Rev Arg Prod Anim 28 (Supl. 1):149-150.
5. Cavestany D, Fernández D, Salazar E, Sanchez A, Leyton L, Crespi D. (2008). Determinación de niveles de progesterona en sangre luego de la administración parenteral de progesterona en vacas Holstein ovariectomizadas. XXXVI Jornadas Uruguayas de Buiatría, Paysandú, Uruguay.
6. Chebel RC, Al-Hassan MJ, Fricke PM, Santos JE, Lima JR, Martel CA, Stevenson JS, García R, Ax RL. (2010). Supplementation of progesterone via controlled internal drug release inserts during ovulation synchronization protocols in lactating dairy cows. J Dairy Sci 93:922-931.
7. Colazo MG, Martínez MF, Whittaker PR, Kastelic JP, Mapletoft RJ. (2002). Estradiol cypionate (ECP) in CIDR-B-based programs for fixed time AI in beef heifers. Theriogenology 57:371 abstr.
8. Colazo MG, Kastelic JP, Mapletoft RJ. (2003). Effects of estradiol cypionate (ECP) on ovarian follicular dynamics, synchrony of ovulation, and fertility in CIDR-based, fixed-time AI programs in beef heifers. Theriogenology 60:855–865.
9. Cutaia L, Chesta P, Moreno D, Bó GA. (2003). Efecto del momento de la aplicación de benzoato de estradiol sobre la sincronía, el tiempo de ovulación y los porcentajes de preñez en vacas tratadas con un dispositivo DIB y PGF. V Simposio Int Reprod Anim. Córdoba. Argentina. Abstr. 386
10. Edmonson AJ, Lean IJ, Weaver LD, Farver T, Webster G. (1989). A body condition scoring chart for Holstein dairy cows. J Dairy Sci 72:68-78.
11. Frank T, Anderson KL, Smith AR, Whitmore HL, Gustafsson BK. (1983). Phagocytosis in the uterus: a review. Theriogenology 20:103-111.
12. Haughian JM, Sartori R, Guenther JN, Gumen A, Wiltbank MC. (2002). Extending the postpartum anovulatory period in dairy cattle with estradiol cypionate. J Dairy Sci 85:3238-3249.
13. Hawk HW. (1975). Hormonal control of changes in the direction of uterine contractions in the estrous ewe. Biol Reprod 12:423-430.
14. Krzymowski T, Stefanczyk-Krzymowska S. (2004). The oestrous cycle and early pregnancy - a new concept of local endocrine regulation. Vet J:168, 285-296.
15. Lammoglia MA, Short RE, Bellows SE, Bellows RA, MacNeil MD, Hafs HD. (1998). Induced and synchronized estrus in cattle: dose titration of estradiol benzoate in peripubertal heifers and postpartum cows after treatment with an intravaginal progesteronereleasing insert and prostaglandin F2α. J Anim Sci 76:1662-1670.
16. Littell RC, Henry PR, Ammermann CB. (1998). Statistical analysis of repeated measures data using SAS procedures. J Anim Sci 76:1216:1231.
17. Lopez H, Satter LD, Wiltbank MC. (2004). Relationship between level of milk production and estrous behavior of lactating dairy cows. Anim Reprod Sci 81:209-223.
18. Lucy MC. (2001). Reproductive loss in high-producing dairy cattle: where will it end? J Dairy Sci 84:1277-1293.
19. Lyimo ZC, Nielen M, Ouweltjes W, Kruip TAM, van Eerdenburg FJCM. (2000). Relationships among estradiol, cortisol and intensity of estrous behavior in dairy cattle. Theriogenology 53:1783-1795.
20. Macmillan KL. (2010). Recent advances in the synchronization of estrus and ovulation in dairy cows. J Reprod Dev 56:S42-S47.
21. Mann GE, Lamming GE. (2000). The role of suboptimal preovulatory oestradiol secretion in the aetiology of premature luteolysis during the short oestrous cycle in the cow. Anim Reprod Sci 64:171- 180.
22. Martinez MF, Kastelic JP, Bo GA, Caccia M, Mapletoft RJ. (2005). Effects of oestradiol and some of its esters on gonadotrophin release and ovarian follicular dynamics in CIDR-treated beef cattle. Anim Reprod Sci 86:37-52.
23. Mattews JN, Altman DG, Campbell NJ, Royston P. (1990). Analysis of serial measurements in medical research. BMJ 27;300:230-235.
24. Meikle A, Sahlin L, Ferraris A, Masironi B, Blanc JE, Rodríguez-Irazoqui M, Rodríguez-Piñón M, Kindahl H, Forsberg M. (2001). Endometrial mRNA expression of oestrogen receptor α, progesterone receptor and insulin-like growth factor-I (IGF-I) throughout the bovine oestrous cycle. Anim Reprod Sci 68:45–56.
25. Murugavel K, Yántiz JL, Santolaria P, López-Béjar M, López-Gatius F. (2003). Luteal activity at the onset of a timed insemination protocol affects reproductive outcome in early postpartum dairy cows. Theriogenology 60:583-593.
26. Pursley JR, Mee MO, Wiltbank MC. (1995). Synchronization of ovulation in dairy cows using PGF 2 Alfa and GnRH. Theriogenology 44:915-923.
27. Rogan D, Martinez MF, Bó GA, Chesta P, Feresin F, Mapletoft RJ. (2007). Progesterone release patterns from Cue-Mate in comparison to other intravaginal progesterone-releasing devices in lactating dairy cows. Reprod Fert Dev 19:126-127.
28. Sangsritavong S, Combs DK, Sartori R, Wiltbank MC. (2002). High feed intake increases blood flow and metabolism of progesterone and estradiol-17β in dairy cattle. J Dairy Sci 85:2831-2842.
29. Sartori R, Rosa GJ, Wiltbank MC. (2002). Ovarian structures and circulating steroids in heifers and lactating cows in summer and lactating cows and dry cows in winter. J Dairy Sci 85:2813-2822.
30. Souza AH, Cunha AP, Caraviello, Wiltbank MC. (2005). Profiles of circulating estradiol-17β after different estrogen treatments in lactating dairy cows. Anim Reprod Sci 2-4:224-232.
31. Stevenson JS, Pursley JR, Garverick HA, Fricke PM, Kesler DJ, Ottobre JS, Wiltbank MC. (2006). Treatment of cycling and noncycling lactating dairy cows with progesterone during Ovsynch. J Dairy Sci 89:2567-2578.
32. Vynckier L, Debackere M, de Kruif A, Coryn M. (1990). Plasma estradiol-17β concentrations in the cow during induced estrus and after injection of estradiol- 17β benzoate and estradiol-17β cyprionate-a preliminary study. J Vet Pharmacol Ther 13:36-42.
33. Wiltbank MC, Lopez H, Sartori R, Sangsritavong S, Gumen A. (2006). Changes in reproductive physiology of lactating dairy cows due to elevated steroid metabolism. Theriogenology 65:17-29.
34. Wolfenson D, Inbar G, Roth Z, Kaim M, Bloch A, Braw-Tal R. (2004). Follicular dynamics and concentrations of steroids and gonadotropins in lactating cows and nulliparous heifers. Theriogenology 62:1042-1055.

