










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
Related links
Share

 Permalink
PermalinkRevista Médica del Uruguay
On-line version ISSN 1688-0390
Rev. Méd. Urug. vol.30 no.2 Montevideo June 2014
Avances en el estudio de la neurobiología de la depresión: rol de la hormona concentradora de melanina
Pablo Torterolo*, Cecilia Scorza†, Jessika Urbanavicius‡, Andrea Devera§, Luciana Benedetto¶, Claudia Pascovich¶, Patricia Lagos**, Michael H. Chase††, Jaime Monti‡‡
Resumen
Introducción: la depresión mayor (DM) es una enfermedad psiquiátrica frecuente, con importante morbilidad y una relación estrecha con el suicidio. Objetivo: hacer una puesta a punto de los avances en el estudio de la neurobiología de la DM, enfocándonos en el posible rol de la hormona concentradora de melanina (MCH) en esta patología. Metodología: revisión de la bibliografía con énfasis en nuestros propios trabajos originales. Resultados: la MCH es un neuromodulador peptídico sintetizado por neuronas del hipotálamo. Las neuronas MCHérgicas envían proyecciones hacia diversas regiones del sistema nervioso central, incluyendo las áreas vinculadas con la regulación de la vigilia y del sueño, así como a diversas estructuras del sistema límbico que participan en la regulación del humor. Aunque numerosos estudios han relacionado el sistema MCHérgico con el control de la homeostasis energética, hallazgos recientes han permitido señalar un rol de este sistema en los mecanismos de generación del sueño. A su vez, una convergencia de datos provenientes de diversos estudios sugiere que la MCH estaría involucrada en la fisiopatología de la DM. Nuestros propios estudios preclínicos tienden a indicar que la MCH promueve la generación del sueño REM y un estado tipo depresivo. Ambos efectos estarían siendo mediados a través de la modulación de la actividad de las neuronas serotoninérgicas del núcleo dorsal del rafe. Conclusiones: estudios preclínicos sugieren un rol protagónico del sistema MCHérgico en la fisiopatología de la depresión. Resta confirmar si esta afirmación es cierta en pacientes con DM.
Palabras clave: NEUROBIOLOGÍA DEPRESIÓN TRASTORNO DEPRESIVO MAYOR HORMONA CONCENTRADORA DE MELANINA
Keywords: NEUROBIOLOGY DEPRESSION DEPRESSIVE DISORDER MAJOR MELANIN CONCENTRATING HORMONE
* Profesor Agregado del Departamento de Fisiología, Facultad de Medicina, Universidad de la República, Uruguay.
† Investigadora Jefa del Departamento de Neurofarmacología Experimental, Instituto de Investigaciones Biológicas Clemente Estable, (IIBCE) Uruguay.
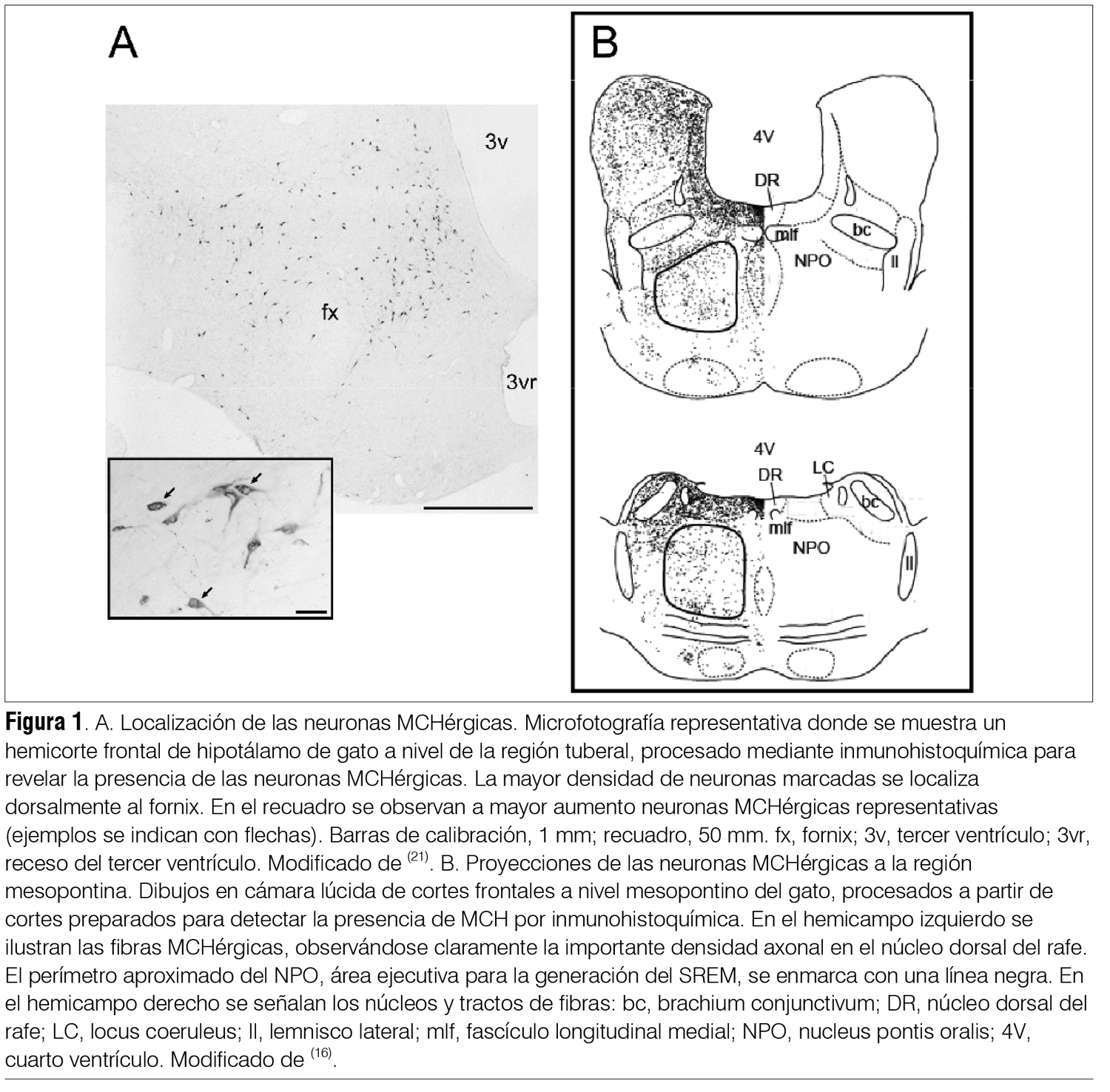
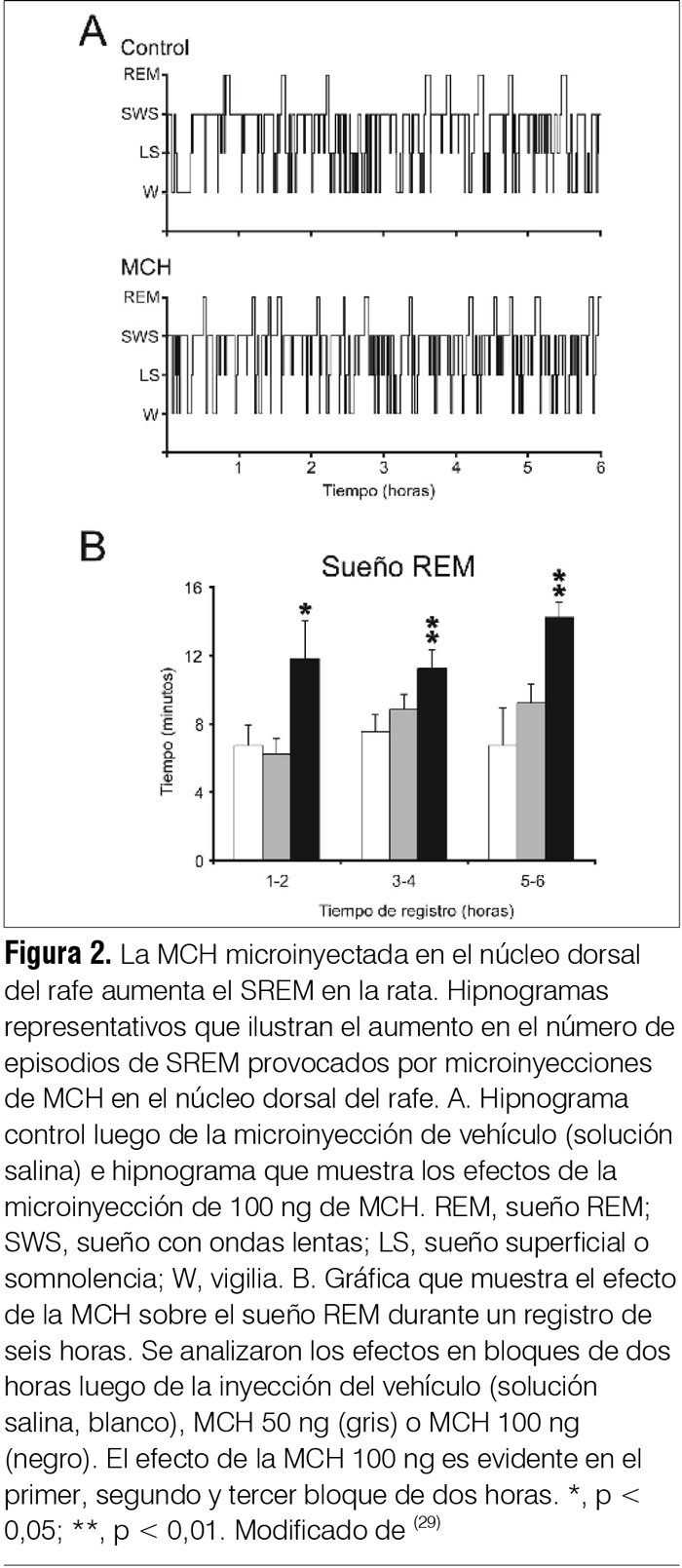
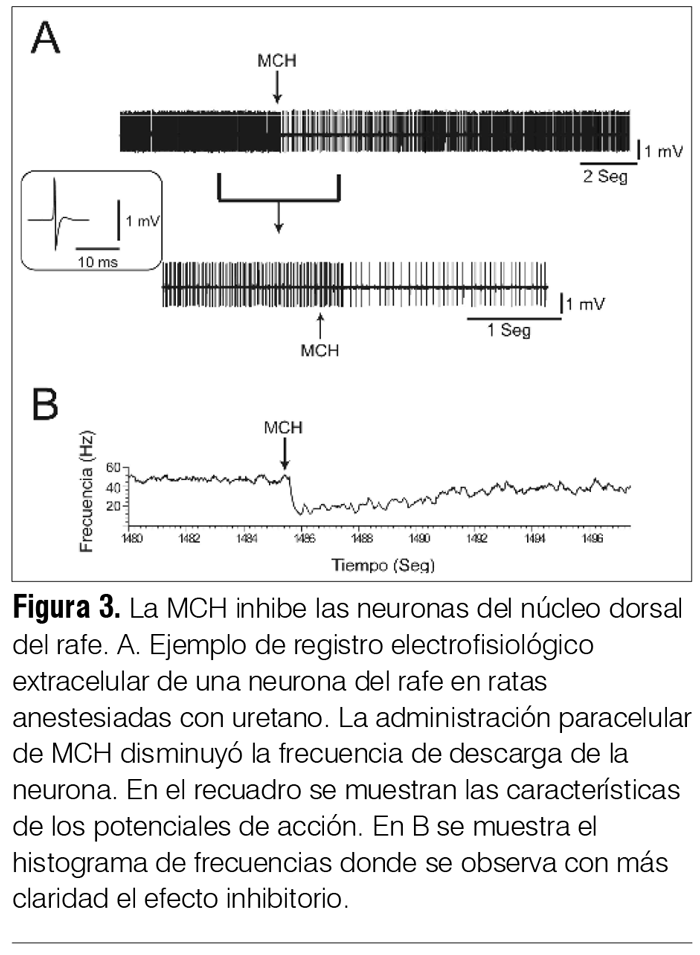
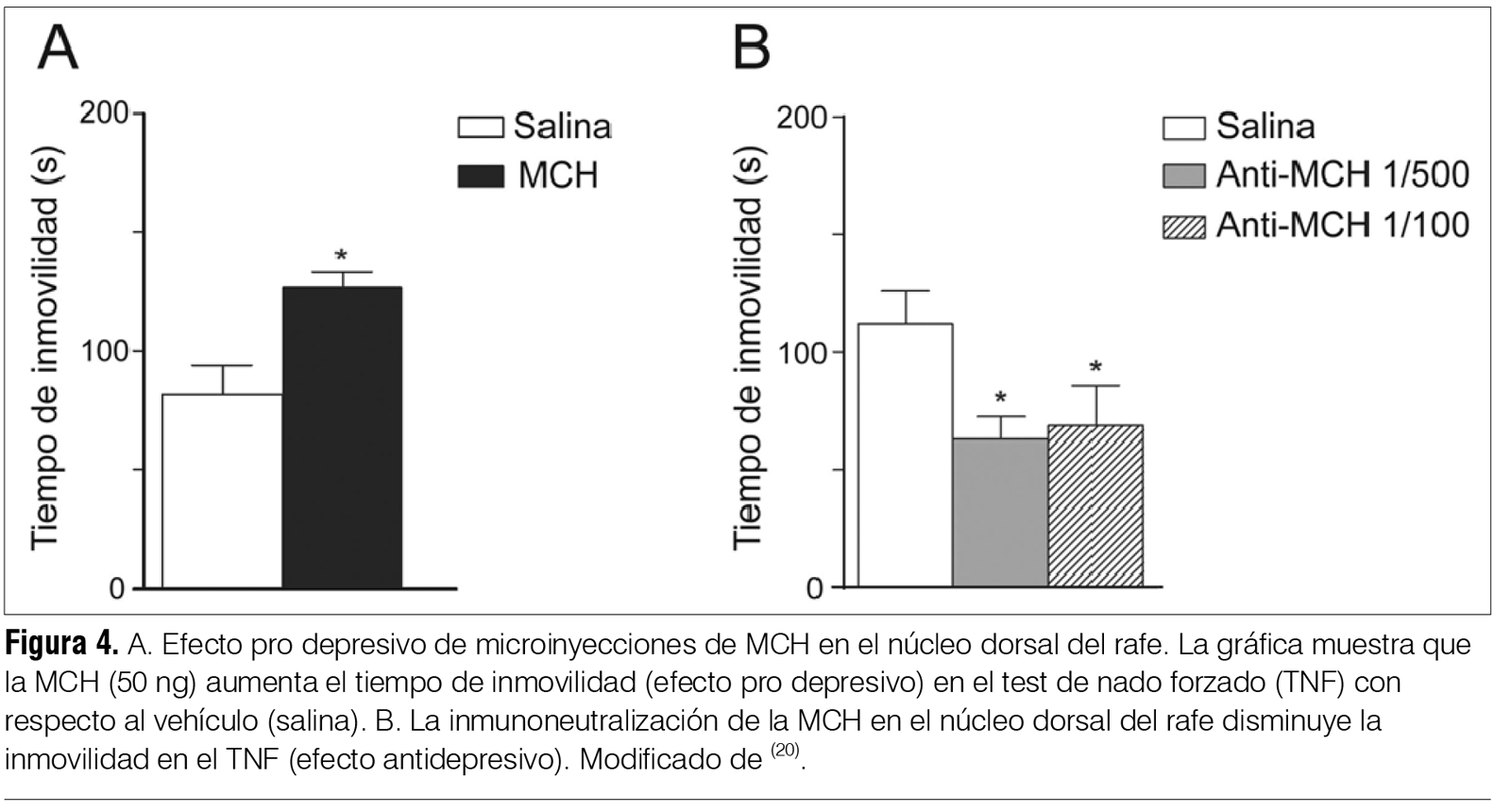
‡ Investigadora del Laboratorio de Biología Celular, IIBCE, Uruguay. § Ex Asistente del Departamento de Fisiología. Asistente de Neonatología, Facultad de Medicina, Universidad de la República, Uruguay. ¶ Asistente, Departamento de Fisiología, Facultad de Medicina, Universidad de la República, Uruguay. ** Prof. Adjunto, Departamento de Fisiología, Facultad de Medicina, Universidad de la República, Uruguay. †† WebSciences International and University of California Los Angeles (UCLA) School of Medicine, USA. Prof. Emérito de UCLA. Prof. Ad Honorem de la Facultad de Medicina, Universidad de la República, Uruguay. ‡‡ Prof. Ex Director del Departamento de Farmacología y Terapéutica, Facultad de Medicina, Universidad de la República, Uruguay. Correspondencia: Dr. Pablo Torterolo. Departamento de Fisiología, Facultad de Medicina, Universidad de la República. Avenida General Flores 2125, CP 11800, Montevideo, Uruguay. Correo electrónico: ptortero@fmed.edu.uy Agradecimientos. El proyecto de investigación del que forma parte este trabajo está siendo financiado por ANII (FCE-1-2011-1-5997) y PEDECIBA. Recibido: 2/12/13 Aceptado: 24/4/14 Introducción La depresión mayor (DM) integra síntomas de tristeza o desesperanza, culpa, irritabilidad y anhedonia, así como síntomas acompañantes tales como cambios del peso corporal, insomnio o hipersomnia, fatiga, falta de concentración e ideas recurrentes de muerte(1). La DM es una de las enfermedades psiquiátricas más comunes en la población general. Estudios epidemiológicos indican que la prevalencia para generar DM es de 8% a 12% en hombres y de 20% a 26% en mujeres. Las repercusiones sanitarias y sociales de la DM son considerables, siendo en los países desarrollados la segunda causa de invalidez. La importancia de la DM aumenta debido a su estrecha relación con el suicidio; se considera que esta patología es la causa de 64% de los suicidios y que 15% de pacientes con DM severa comete suicidio. Uruguay es el país sudamericano (y segundo en Latinoamérica) con mayor prevalencia de suicidios. Según datos del Ministerio de Salud Pública, en el 2002 la tasa de suicidios fue de 21.7 cada 100.000 habitantes y en 2010 hubo 559 muertes por esta causa. El suicidio es la causa del 12% a 15% de las muertes en jóvenes de 25 a 34 años(2,3). Etiología y patogenia de la depresión. Rol del sistema serotoninérgico A pesar de los avances en la neurobiología de la depresión(4-7) todavía se tienen escasos conocimientos sobre su etiopatogenia. Diversos estudios han mostrado un factor genético en la vulnerabilidad a padecer depresión. Si bien la DM está relacionada fuertemente con diversos eventos estresantes, el estrés per se no es una condición suficiente para causar la patología(4,6). Otros factores, como traumas emocionales en etapas tempranas del desarrollo, han sido también implicados en el origen de la enfermedad(4-6). Múltiples evidencias involucran al sistema de neurotransmisión serotoninérgico en la fisiopatología de la DM. El sistema serotoninérgico comprende uno de los sistemas neuroquímicos más distribuidos en el sistema nervioso central (SNC), localizándose la mayoría de los somas de las neuronas serotoninérgicas en el núcleo dorsal del rafe (NDR)(8). Mediante proyecciones a múltiples regiones del SNC, las neuronas serotoninérgicas desempeñan un rol importante no solo en la regulación de los estados emocionales sino también en diversas funciones entre las que se destaca el control del ciclo sueño-vigilia. Se ha observado que un importante porcentaje de pacientes con DM o suicidas, o ambos, presentan una disminución de los niveles de serotonina o sus metabolitos, o ambos, en la orina y en el líquido cefalorraquídeo (LCR)(4-6). La presencia de distintos polimorfismos de transportadores, enzimas o receptores relacionados con la serotonina también se han relacionado con la DM, con intentos de autoeliminación o suicidio(9). Por último, los fármacos inhibidores selectivos de la recaptación de serotonina (ISRS) son antidepresivos(5). Se han encontrado diferencias en marcadores serotoninérgicos en el NDR de pacientes depresivos-suicidas comparados con individuos control; por lo tanto, las neuronas serotoninérgicas del NDR constituyen un sitio potencial de disfunción en la neurobiología de la DM y un blanco importante de los agentes antidepresivos. Otras líneas de trabajo han mostrado en ciertos pacientes depresivos anormalidades en el volumen, flujo sanguíneo, metabolismo o patrones de activación de estructuras cerebrales tales como la corteza prefrontal, la corteza cingulada, la amígdala y el hipocampo; regiones que reciben aferencias serotoninérgicas(4-6). También se ha observado en subgrupos de pacientes depresivos una hiperactivación del eje hipotálamo-hipofisario-adrenal; además, la administración sistémica de dexametasona (glucocorticoide sintético) no suprime la secreción de cortisol como ocurre en individuos normales(4,6). Un problema relacionado con el tratamiento utilizando fármacos antidepresivos es la prolongada latencia en la aparición de sus efectos terapéuticos a pesar de que dichos fármacos provocan un aumento inmediato en los niveles extracelulares de monoaminas. Este hecho sugiere que el efecto bioquímico no determina el efecto terapéutico, sino que este sería producido por cambios neurobiológicos más lentos, entre los que se han sugerido la desensibilización de receptores serotoninérgicos, la modulación de vías intracelulares y la expresión de genes de factores de crecimiento como el brain-derived neurotrophic factor (BNDF) y la regulación de la neurogénesis posnatal(4,6). Depresión y sueño REM Los pacientes con DM se caracterizan por presentar una disminución de la latencia para la aparición del primer período de sueño con movimientos oculares rápidos o rapid eye movements o REM (SREM, también llamado sueño paradójico), siendo este uno de los signos biológicos más robustos y específicos de la DM(7). A su vez, presentan un aumento del tiempo total de SREM, un incremento de la duración del primer período de SREM y de la densidad de los movimientos oculares rápidos durante esta etapa. El hecho de que la mayoría de los fármacos antidepresivos disminuyan o supriman el SREM, y que la privación total de sueño o selectiva de SREM sea eficaz en el tratamiento de la DM, subraya la relación entre esta patología y el SREM(7). Se puede considerar entonces que los pacientes con DM tienen un aumento de la “presión” para generar SREM, o que la generación del SREM está promovida o facilitada en estos pacientes. Alteraciones similares del SREM también se observan en modelos animales de DM. Los efectos de los fármacos antidepresivos han sido muy estudiados en animales mediante el test de nado forzado (TNF). Luego de unos minutos en dicha situación de estrés, los animales entran en un estado que ha sido llamado de desesperanza, en el que dejan de nadar y adoptan una postura de inmovilidad. Los fármacos con acción antidepresiva disminuyen el tiempo de inmovilidad mientras que aumentan el tiempo de nado y de escalamiento (intentos de trepar por las paredes del recipiente). Dichas respuestas son independientes de los cambios que pudieran generar los fármacos sobre la actividad locomotora. Se destaca que en ratas la privación de sueño por 24 horas genera un aumento en el tiempo de nado que se interpreta como un efecto antidepresivo; este efecto se ha vinculado con un aumento en la actividad serotoninérgica(10). Además, la privación de sueño potencia el efecto del tratamiento farmacológico con antidepresivos. La relación entre el SREM y la DM parecería depender, por lo menos en parte, de las neuronas serotoninérgicas del NDR. Estas intervendrían tanto en la generación del SREM como en la fisiopatología de la DM(7). Sistema serotoninérgico: rol en el sueño REM Las neuronas serotoninérgicas del NDR presentan una descarga tónica durante la vigilia, decrecen su actividad durante el sueño con ondas lentas (SL, también llamado sueño lento o no REM) y se inactivan casi totalmente durante el SREM (neuronas REM-off)(8). La disminución de la liberación de serotonina en diversas áreas cerebrales durante el SREM se correlaciona con los hallazgos electrofisiológicos. El neurotransmisor ácido gamma-aminobutírico (GABA) y las neuronas GABAérgicas del NDR estarían involucradas en la inhibición de las neuronas serotoninérgicas durante el SREM y de la consiguiente promoción de dicho estado de comportamiento(11). Por otro lado, la activación experimental de las neuronas serotoninérgicas del NDR bloquea la generación del SREM. De esta serie de evidencias se puede concluir que la generación del SREM dependería en forma considerable de la disminución del tono serotoninérgico, fenómeno que ha llevado a plantear que estas neuronas sean consideradas “permisivas” para la generación del mismo. MCH: generalidades La MCH es un péptido cíclico de 19 aminoácidos que fue inicialmente caracterizado como un factor circulante que mediaba cambios de color en los peces teleósteos(12-14). Posteriormente se identificó como neurotransmisor/neuromodulador en mamíferos, incluyendo a los seres humanos. La función biológica de la MCH es mediada por dos receptores acoplados a la proteína-G, MCHR-1 y MCHR-2. Es interesante destacar que el gen del MCHR-2 es un pseudogen en roedores, pero es funcional en carnívoros y primates, incluyendo el ser humano. Se ha determinado que el MCH-R1 activa las proteínas Gi y Gq e inhibe corrientes de Ca++. También se ha demostrado que la MCH tiene un rol principalmente inhibitorio tanto a nivel presináptico, donde disminuye la liberación de GABA y glutamato, como a nivel postsináptico(12-14). La MCH se sintetiza en neuronas cuyos somas están localizados en el hipotálamo posterolateral y en la zona incerto-hipotalámica (figura 1A); estas neuronas proyectan a diversas regiones del SNC(14-16). La MCH y el control de la homeostasis energética Este sistema neuronal estuvo tradicionalmente relacionado con el control de la homeostasis energética, es decir, de la ingesta y de la actividad metabólica(12). Al respecto, la infusión crónica de un agonista sintético del receptor MCHR-1 induce obesidad en ratones, acompañada de hiperfagia, reducción de la temperatura corporal y una estimulación de la actividad lipogénica en el hígado y en el tejido adiposo blanco. A su vez, animales genéticamente modificados con sobreexpresión de MCH presentan obesidad, mientras que animales que carecen de MCH presentan hipofagia y bajo peso corporal. Estas evidencias sugieren que aumentando la ingesta y favoreciendo el anabolismo, la MCH promueve la conservación de la energía corporal. El sistema MCHérgico promueve el sueño Una de las principales funciones del sueño es la conservación de la energía. Por lo tanto, es esperable que las neuronas MCHérgicas, que son críticas en el control de la homeostasis energética, estén involucradas en el sueño. En efecto, hace diez años aparecieron las primeras evidencias experimentales indicando que el sistema MCHérgico estaría involucrado en los mecanismos de generación del sueño(17). La MCH, a través de la regulación de los sistemas activadores y somnogénicos(18), promovería el sueño, especialmente el SREM(13,14). Los principales argumentos experimentales que sostienen un rol del sistema MCHérgico como promotor del sueño se detallan a continuación. Anatomía del sistema MCHérgico. Relación con los sistemas activadores Estudios clásicos han relacionado la región posterolateral del hipotálamo, donde se encuentran las neuronas MCHérgicas, con el control del ciclo sueño-vigilia(18). A su vez, estas neuronas presentan una estrecha relación anatómica con las neuronas hipocretinérgicas (también conocidas como orexinérgicas), cuyos somas también se encuentran en esta región(15). Estas neuronas son esenciales para el mantenimiento de la vigilia y su degeneración está en la base patogénica de la narcolepsia(19). Las neuronas MCHérgicas proyectan densamente a regiones activadoras y somnogénicas(14). Mediante la utilización de marcadores retrógrados hemos caracterizado las proyecciones de las neuronas MCHérgicas al núcleo pontis oralis (NPO), un área ejecutiva para la generación de SREM(16). También existe una alta densidad de fibras en regiones activadoras tales como el NDR y el locus coeruleus (región con densa población de neuronas noradrenérgicas)(20,21). En la figura 1B se exhibe la distribución de las fibras MCHérgicas en la región mesopontina, destacándose la importante inervación del NDR. Regiones del sistema límbico, que participan en el control de los estados emocionales, también reciben una alta densidad de aferencias MCHérgicas y expresan receptores para la misma. También hemos documentado que los tanicitos del NDR del gato tienen inmunorreactividad para la MCH. Los tanicitos son células especializadas que presentan un soma ependimario y un largo proceso basal que penetra en el parénquima nervioso; estas células absorben sustancias presentes en el LCR y las pueden transportar hacia el parénquima (21). Este dato y la documentada presencia de MCH en el LCR de ratas, ovejas y humanos(22-24), sugieren que el sistema MCHérgico podría regular la actividad de los núcleos del rafe mesopontinos a través de una vía neurohumoral (por conducción de volumen utilizando el sistema ventricular), complementando la vía neural directa. Se ha identificado una distribución amplia de los receptores MCHR-1 en el SNC de la rata, que coincide con la distribución de las fibras MCHérgicas. Resultados preliminares de nuestro grupo sugieren que las neuronas serotoninérgicas del NDR expresan receptores para MCH. Las neuronas MCHérgicas se activan durante el sueño Utilizando la proteína Fos como índice de actividad neuronal se ha demostrado que las neuronas MCHérgicas se activan durante el SREM de la rata(25). Hassani y colaboradores, en el año 2009, registraron las neuronas MCHérgicas; estas neuronas presentan muy baja frecuencia de descarga durante la vigilia, aumentan levemente su frecuencia de descarga durante el SL y se activan al máximo durante el SREM(26). Titulación de los niveles de MCH durante la vigilia y el sueño La concentración de MCH en el LCR de la rata aumenta durante la etapa de luz, cuando el animal está preferentemente dormido, mientras que disminuye durante la noche, período en el que está mayoritariamente despierto(23). A su vez, mediante microdiálisis se ha demostrado que la liberación de MCH en la amígdala de pacientes con epilepsia resistente al tratamiento farmacológico es mínima durante la vigilia activa con interacción social, se incrementa luego de ingerir alimentos (etapa consumatoria), mientras que es máxima al comienzo del sueño(27). Estudios en animales genéticamente modificados Estudios en ratones knock-out del receptor MCHR-1 así como para la MCH (supresión del gen que los codifica) indican que dichos animales poseen una alteración de la arquitectura del sueño(14). Los animales que carecen de MCH presentan menos SREM, lo que se hace especialmente evidente en condiciones de balance energético negativo. Administración de MCH o antagonistas del receptor MCHR-1 La administración intracerebroventricular de MCH en la rata produce un marcado aumento del SREM y un moderado aumento del SL(25). A su vez, la administración sistémica de antagonistas de los receptores MCHR-1 en la misma especie disminuye el SL y el SREM e incrementa el tiempo de vigilia(28). Como se observa en la figura 2, microinyecciones de MCH en el NDR de la rata facilitan la generación de SREM(29). Por el contrario, la inmunoneutralización de la MCH endógena produce el efecto contrario(30). Estudios preliminares en gatos (donde estarían activos los dos tipos de receptores de MCH) también han mostrado que microinyecciones de MCH en el NDR producen un aumento del SL o del SREM, dependiendo del lugar exacto de las microinyecciones. La MCH también promueve el SREM actuando tanto a nivel del cerebro basal anterior de la rata como del NPO del gato(16,31); estas áreas se encuentran relacionadas con la generación de este estado de comportamiento. Por el contrario, la administración de MCH en el área preóptica ventrolateral en la rata, área implicada en la generación del SL, favorece su generación(32). La activación experimental de las neuronas MCHérgicas induce el sueño Konadhode y colaboradores, en el año 2013, utilizaron la técnica de optogenética para estimular específicamente las neuronas MCHérgicas en ratones(33). Los autores insertaron el gen para el canal catiónico fotosensible channelrhodopsine-2 en las neuronas MCHérgicas y posteriormente fueron capaces de estimularlas de forma específica con un estímulo luminoso. El protocolo de estimulación utilizado indujo una disminución de la latencia al sueño, reducción de la duración del tiempo promedio de vigilia, así como un incremento del tiempo total de SL y SREM durante la noche; durante el día aumentó la profundidad del sueño. Los autores sostienen que la actividad MCHérgica es capaz de contrarrestar la actividad combinada de los sistemas activadores y, por lo tanto, agonistas MCHérgicos podrían ser potencialmente útiles para el tratamiento del insomnio. Efecto de la MCH sobre neuronas serotoninérgicas En registros extracelulares in vivo hemos observado que la aplicación intracerebroventricular o paracelular de MCH inhibe la mayoría de las neuronas serotoninérgicas (reconocidas por sus características electrofisiológicas y farmacológicas) del NDR y del núcleo mediano del rafe. En la figura 3 se muestra un ejemplo del efecto inhibitorio de la aplicación paracelular de MCH sobre neuronas del NDR. Asimismo, la aplicación de bajas concentraciones de MCH en el NDR disminuye los niveles extracelulares de serotonina en esta misma región(34). Rol del sistema MCHérgico en la fisiopatología de la depresión Como se mencionó anteriormente, la importante densidad de fibras MCHérgicas en el NDR, la presencia de MCHR-1 en las neuronas serotoninérgicas de este núcleo, así como las importantes proyecciones al sistema límbico, sugieren un rol de la MCH en el control de los estados emocionales. A su vez, datos indirectos, como que la MCH facilita el SREM (y el SREM está aumentado en la DM) y estimula el eje hipotálamo-hipofisario-adrenal (que está sobreactivado en la DM) también sugieren que una hiperactividad del sistema MCHérgico podría estar relacionada con algunos aspectos de la DM. Borowsky y colaboradores, en el año 2002, demostraron que un antagonista de los receptores MCHR-1, SNAP-7941, posee un perfil antidepresivo y ansiolítico al ser evaluado en modelos animales(35). Algunos autores, utilizando otros antagonistas así como diversos modelos animales de depresión, obtuvieron resultados similares(36,37). La importancia del sistema MCHérgico en la DM es enfatizada en un estudio reciente que propone que un aumento de la expresión de la preproMCH y una consecuente downregulation de los receptores MCHérgicos podría ser un biomarcador de la severidad de los comportamientos depresivos(38). Utilizando el TNF en ratas exploramos si la aplicación de MCH en el NDR generaba comportamientos tipo depresivos(20). En esta serie de experimentos hemos encontrado que la MCH microinyectada en el NDR induce una respuesta pro depresiva aumentando significativamente el tiempo de inmovilidad (efecto opuesto al antidepresivo) en el TNF; este efecto se muestra en la figura 4A. Esta respuesta se bloquea si los animales son previamente tratados de forma sistémica con fluoxetina, antidepresivo del grupo de los ISRS. Por otra parte, como se muestra en la figura 1B, la inmunoneutralización de la MCH endógena produce una respuesta antidepresiva, disminuyendo significativamente el tiempo de inmovilidad(20). Esta respuesta estuvo acompañada de un aumento en el tiempo de nado; ambos cambios se asocian a un aumento en la neurotransmisión serotoninérgica(20). El efecto pro depresivo podría relacionarse con la inhibición de la actividad serotoninérgica en el NDR inducida por la MCH, sugerida por estudios electrofisiológicos y por microdiálisis (especificado anteriormente). Por último, estudios recientes han demostrado que el núcleo mediano del rafe también participa en el efecto pro depresivo inducido por MCH(39). Los trabajos de nuestro grupo, en concordancia con los de otros autores(36,37), sugieren que la DM podría relacionarse con un aumento en la actividad de las neuronas MCHérgicas. Si esta hipótesis fuese correcta, sería de esperar que los antidepresivos disminuyeran la actividad de estas neuronas. De manera interesante se ha observado que el tratamiento agudo con escitalopram (antidepresivo del grupo de los ISRS) inhibe tanto el rebote de SREM luego de una privación de sueño como la actividad de las neuronas MCHérgicas(40). Ambos efectos irían a favor de la hipótesis planteada. Dado que las neuronas MCHérgicas regulan la homeostasis energética es esperable que durante la DM esta función esté alterada; de hecho, es característico de esta patología las alteraciones en el peso corporal(1). A su vez, estudios preclínicos han demostrado que los antagonistas de los MCHR-1 no solo tienen efectos antidepresivos, sino que también tienen un fuerte efecto antiobesidad(36,37). Por último, dado que el número de las neuronas MCHérgicas aumenta en el posparto en los circuitos del área preóptica que controlan el comportamiento maternal, y que la MCH regula este comportamiento(41), también sería importante conocer si la MCH interviene en los desajustes emocionales que ocurren en 75% a 80% de las madres entre tres a cinco días luego del parto. Conclusiones y perspectivas Estudios preclínicos sugieren un rol protagónico del sistema MCHérgico en el control del SREM y de la depresión. Nuestra hipótesis de trabajo se basa en que un aumento de la actividad de las neuronas MCHérgicas podría estar en la base patogénica de la depresión. De acuerdo con esta hipótesis, sería importante evaluar si pacientes depresivos presentan un aumento de los niveles de MCH a nivel del LCR. Por último, trabajos preclínicos han demostrado que los antagonistas de los receptores MCHR-1 tienen un enorme potencial como fármacos antidepresivos. Su eficiencia, el rápido inicio de sus efectos así como la aparente ausencia de efectos severos en el SNC, sugieren la potencialidad de estos fármacos(36,37). Abstract Introduction: major depression disorder (MDD) is a common psychiatric condition, it has high morbility rates and is closely related to suicide. Objective: to provide an update in the study of the neurobiology of depression, focusing on the potential role of the melanin-concentrating hormone (MCH) in this condition. Method: a bibliographical review with an emphasis on our own original studies. Results: the melanin-concentrating hormone is a peptide neuromodulator syhthetized by neurones in the hypothalamus. MCHergic neurons send projection towards different areas in the central nervous system, including areas associated to the regulation of the sleep-wake cycle, as well as different structures in the limbic system that take part in the regulation of mood. In spite of several studies having proved the MCHergic system with the control of energetic homeostasis, recent findings have enabled to identify a role for this system in the sleep generator mechanisms. Similarly, data arising from several studies suggests that MCH would be involved in the major depression disorder. Our own preclinical studies tend to pint out the MCH promotes the generation of REM sleep and a type of depression. Apparently both effects would be mediated through the modulation of the activity on the serotoninergic neurons in the dorsal raphe nucleus. Conclusions: paraclinical studies suggest the leading role of the MCHergic system in the pathophysiology of depression. It is to be proved still, whether this affirmation is true for patients with major depression disorder. Resumo Introdução: a depressão maior (DM) é uma enfermidade psiquiátrica frequente, com morbidade considerável e estreitamente relacionada com o suicídio. Objetivo: fazer uma atualização dos avanços no estudo da neurobiologia da DM, focando no possível papel do hormônio concentrador melanina (MCH) nesta patologia. Metodologia: revisão da bibliografia com ênfase em nossos trabalhos originais. Resultados: o MCH é um neuromodulador peptídico sintetizado pelos neurônios do hipotálamo. Os neurônios MCHérgicos enviam projeções a diversas regiões del sistema nervoso central, incluindo as áreas vinculadas com a regulação da vigília e do sono, bem como a diversas estruturas do sistema límbico que participam na regulação do humor. Embora numerosos estudos hajam relacionado o sistema MCHérgico com o controle da homeostase energética, descobrimentos recentes permitiram identificar um papel deste sistema nos mecanismos de geração do sono. Por outro lado, uma convergência de dados provenientes de diversos estudos sugere que o MCH estaria relacionado com a fisiopatologia da DM. Nossos estudos preclínicos tendem a indicar que o MCH promove a geração do sono REM e um estado tipo depressivo. Ambos efeitos estariam sendo mediados pela modulação da atividade dos neurônios serotoninérgicos do núcleo dorsal do rafe. Conclusões: estudos preclínicos sugerem um protagonismo do sistema MCHérgico na fisiopatologia da depressão. É necessário confirmar se esta afirmação é correta em pacientes com DM. Bibliografía 1. American Psychiatric Association. Diagnostic and statistical manual of mental disorders: DSM-IV. 4 ed. Washington (DC): APA, 1994. 2. Montalbán A. El suicidio: la urgencia de un grave problema. Rev Méd Urug 2004; 20(2): 91. 3. Lucero Abreu R. Suicidios en Uruguay: su relación con la economía nacional (1972-1992). Rev Méd Urug 1998; 14(3): 236-47. 4. Saveanu RV, Nemeroff CB. Etiology of depression: genetic and environmental factors. Psychiatr Clin North Am 2012; 35(1):51-71. 5. Willner P, Scheel-Krüger J, Belzung C. The neurobiology of depression and antidepressant action. Neurosci Biobehav Rev 2013; 37(10 Pt 1):2331-71. 6. Palazidou E. The neurobiology of depression. Br Med Bull 2012; 101:127-45. 7. Palagini L, Baglioni C, Ciapparelli A, Gemignani A, Riemann D. REM sleep dysregulation in depression: state of the art. Sleep Med Rev 2013; 17(5):377-90. 8. Monti JM. The structure of the dorsal raphe nucleus and its relevance to the regulation of sleep and wakefulness. Sleep Med Rev 2010; 14(5):307-17. 9. Bondy B, Buettner A, Zill P. Genetics of suicide. Mol Psychiatry 2006; 11(4):336-51. 10. López-Rodríguez F, Kim J, Poland RE. Total sleep deprivation decreases immobility in the forced-swim test. Neuropsychopharmacology 2004; 29(6):1105-11. 11. Torterolo P, Yamuy J, Sampogna S, Morales FR, Chase MH. GABAergic neurons of the cat dorsal raphe nucleus express c-fos during carbachol-induced active sleep. Brain Res 2000; 884(1-2):68-76. 12. Macneil DJ. The role of melanin-concentrating hormone and its receptors in energy homeostasis. Front Endocrinol (Lausanne) 2013; 4:49. 13. Torterolo P, Lagos P, Monti JM. Melanin-concentrating hormone: a new sleep factor? Front Neurol 2011; 2:14. 14. Monti JM, Torterolo P, Lagos P. Melanin-concentrating hormone control of sleep-wake behavior. Sleep Med Rev 2013; 17(4):293-8. 15. Torterolo P, Sampogna S, Morales FR, Chase MH. MCH-containing neurons in the hypothalamus of the cat: searching for a role in the control of sleep and wakefulness. Brain Res 2006; 1119(1):101-14. 16. Torterolo P, Sampogna S, Chase MH. MCHergic projections to the nucleus pontis oralis participate in the control of active (REM) sleep. Brain Res 2009; 1268:76-87. 17. Torterolo P, Rojas M, Sampogna S, Morales FR, Chase MH. MCH-containing neurons and the control of sleep and wakefulness. Sleep 2003; 26:A20. 18. Torterolo P, Vanini G. Nuevos conceptos sobre la generación y el mantenimiento de la vigilia. Rev Neurol 2010; 50(12):747-58. 19. Torterolo P, Vanini G. Importancia de las hipocretinas en al patogenia de la narcolepsia: breve revisión. Rev Méd Urug 2003; 19(1):27-33. 20. Lagos P, Urbanavicius J, Scorza MC, Miraballes R, Torterolo P. Depressive-like profile induced by MCH microinjections into the dorsal raphe nucleus evaluated in the forced swim test. Behav Brain Res 2011; 218(2):259-66. 21. Torterolo P, Lagos P, Sampogna S, Chase MH. Melanin-concentrating hormone (MCH) immunoreactivity in non-neuronal cells within the raphe nuclei and subventricular region of the brainstem of the cat. Brain Res 2008; 1210:163-78. 22. Ungerfeld R, Alzugaray S, Quintela HG, Lagos P, Torterolo P, Bielli A. Melanin concentrating hormone (MCH) in the cerebrospinal fluid of ewes during spontaneous oestrous cycles and ram effect induced follicular phases. Peptides 2011; 32(12):2511-3. 23. Calegare B, Fernandes L, Dias A, Costa A, Lagos P, Torterolo P, et al. Melanin-concentrating hormone levels in the CSF during paradoxical sleep deprivation and rebound: comparison with hypocretin-1 (orexin-A). En prensa 2013. 24. Peyron C, Valentin F, Bayard S, Hanriot L, Bedetti C, Rousset B, et al. Melanin concentrating hormone in central hypersomnia. Sleep Med 2011; 12(8):768-72. 25. Verret L, Goutagny R, Fort P, Cagnon L, Salvert D, Léger L, et al. A role of melanin-concentrating hormone producing neurons in the central regulation of paradoxical sleep. BMC Neurosci 2003; 4:19. 26. Hassani OK, Lee MG, Jones BE. Melanin-concentrating hormone neurons discharge in a reciprocal manner to orexin neurons across the sleep-wake cycle. Proc Natl Acad Sci U S A 2009; 106(7):2418-22. 27. Blouin AM, Fried I, Wilson CL, Staba RJ, Behnke EJ, Lam HA, et al. Human hypocretin and melanin-concentrating hormone levels are linked to emotion and social interaction. Nat Commun 2013; 4:1547. 28. Ahnaou A, Drinkenburg WH, Bouwknecht JA, Alcazar J, Steckler T, Dautzenberg FM. Blocking melanin-concentrating hormone MCH1 receptor affects rat sleep-wake architecture. Eur J Pharmacol 2008; 579(1-3):177-88. 29. Lagos P, Torterolo P, Jantos H, Chase MH, Monti JM. Effects on sleep of melanin-concentrating hormone (MCH) microinjections into the dorsal raphe nucleus. Brain Res 2009; 1265:103-10. 30. Lagos P, Torterolo P, Jantos H, Monti JM. Immunoneutralization of melanin-concentrating hormone (MCH) in the dorsal raphe nucleus: effects on sleep and wakefulness. Brain Res 2011; 1369:112-8. 31. Lagos P, Monti JM, Jantos H, Torterolo P. Microinjection of the melanin-concentrating hormone into the lateral basal forebrain increases REM sleep and reduces wakefulness in the rat. Life Sci 2012; 90(23-24):895-9. 32. Benedetto L, Rodriguez-Servetti Z, Lagos P, D’Almeida V, Monti JM, Torterolo P. Microinjection of melanin concentrating hormone into the lateral preoptic area promotes non-REM sleep in the rat. Peptides 2013; 39:11-5. 33. Konadhode RR, Pelluru D, Blanco-Centurion C, Zayachkivsky A, Liu M, Uhde T, et al. Optogenetic stimulation of MCH neurons increases sleep. J Neurosci 2013; 33(25):10257-63. 34. Urbanavicius J, Lagos P, Torterolo P, Scorza C. Role of melanin concentrating hormone (MCH) on the dorsal raphe nucleus (DRN): its relevance for depression. J Neurochem 2013; 125: 265. 35. Borowsky B, Durkin MM, Ogozalek K, Marzabadi MR, DeLeon J, Lagu B, et al. Antidepressant, anxiolytic and anorectic effects of a melanin-concentrating hormone-1 receptor antagonist. Nat Med 2002; 8(8):825-30. 36. Chung S, Parks GS, Lee C, Civelli O. Recent updates on the melanin-concentrating hormone (MCH) and its receptor system: lessons from MCH1R antagonists. J Mol Neurosci 2011; 43(1):115-21. 37. Shimazaki T, Yoshimizu T, Chaki S. Melanin-concentrating hormone MCH1 receptor antagonists: a potential new approach to the treatment of depression and anxiety disorders. CNS Drugs 2006; 20(10):801-11. 38. García-Fuster MJ, Parks GS, Clinton SM, Watson SJ, Akil H, Civelli O. The melanin-concentrating hormone (MCH) system in an animal model of depression-like behavior. Eur Neuropsychopharmacol 2012; 22(8):607-13. 39. López Hill X, Pascovich C, Urbanavicius J, Torterolo P, Scorza MC. The median raphe nucleus participates in the depressive-like behavior induced by MCH: differences with the dorsal raphe nucleus. Peptides 2013; 50:96-9. 40. Kátai Z, Adori C, Kitka T, Vas S, Kalmár L, Kostyalik D, et al. Acute escitalopram treatment inhibits REM sleep rebound and activation of MCH-expressing neurons in the lateral hypothalamus after long term selective REM sleep deprivation. Psychopharmacology (Berl) 2013; 228(3):439-49. 41. Benedetto L, Pereira M, Ferreirra A, Torterolo P. Melanin-concentrating hormone in the medial preoptic area reduces active components of maternal behavior. Peptides 2014; 58:20-5.


{kind=link}
{kind=link}
{kind=link}
{kind=link}