











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo

 Permalink
Permalink
1. Introduction
One of the main restrictions on horticultural production in Uruguay stems from the damage caused by crop’s pests and the limitations associated with their control. Since the definition and development of the Integrated Pest Management (IPM) concept1, significant progress has been made in its practical application. IPM can be defined as a flexible and holistic system which considers the agroecosystem as an interconnected whole and employs a combination of biological, cultural, genetic, physical and chemical methods to keep populations below economically damaging levels while minimizing disruption to the crop ecosystem and the surrounding environment2. The main objective is to understand, explore and apply different pest control techniques with the aim of achieving a deeper understanding of the importance of integrated pest management and its role in maintaining sustainable ecosystems.
In greenhouses, insects encounter favorable environmental conditions that allow their development almost year-round, along with successive crops which are used as both substrate and food sources. Pests cause significant damage to both the quantity and quality of marketable product, often leading growers to apply numerous chemical insecticide treatments in an attempt to reduce pest populations. However, these efforts frequently fail to achieve satisfactory reductions in pest abundance, resulting in a production system dependent on continuous interventions3. Simultaneously, the use of toxic products poses direct effects on human health, the environment, beneficial insects, the emergence of resistant insect populations and the accumulation of residues on fruit 4)(5) 6. These residues may affect consumer health and limit market access, thereby threatening the stability of production units. Furthermore, the excessive use of pesticides can negatively impact the activity of biological control agents, promote the emergence of secondary pests and disrupt the natural regulatory mechanisms of pest populations7. Pest resurgence often occurs because natural enemies are generally more susceptible to pesticides than the target pests8.
In Uruguay’s horticultural production system, greenhouse tomato crops often coexist with other cultivated vegetables such as pepper, strawberry, beet, lettuce, zucchini, eggplant, and in several cases, with fruit trees (e.g., citrus and peach). The highly heterogeneous landscape and the relatively open structures of greenhouses facilitate interactions with surrounding crops. Additionally, open fields and wild plants act as reservoirs for pests and diseases. However, the surrounding environment is also a significant source of natural enemies, which migrate into and establish themselves in the crops when insecticide use is reduced9. Among the pests that affect greenhouse tomato crops in Uruguay are the western flower thrips Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae), a vector of tomato spotted wilt virus (TSWV); the tomato leafminer Phthorimaea absoluta (Meyrick) (Lepidoptera: Gelechiidae); several species of aphids (Hemiptera: Aphididae), and the whiteflies Trialeurodes vaporariorum (Westwood) and Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae), which are cosmopolitan pests10.
In response to the widespread use of pesticides to control these pests, and the growing regulatory requirements regarding maximum residue limits for fruits and vegetables, there is a growing need to adopt control agents that allow for the production of chemical-free. This trend toward reducing the number of insecticide applications has led to an increased use of biological control programs through the introduction of natural enemies (predators, parasitoids or entomopathogens)11. Biological control in greenhouse environments has proven to be a viable alternative to pesticide use, both environmentally and economically. In this context, the Entomology Unit (Faculty of Agronomy, Uruguay) has established a research group focused on developing the scientific foundations for the biological control of key pests affecting greenhouse tomato crops in Uruguay, with the aim of applying experimentally validated tools to ensure their effectiveness. The IPM recognizes that there is no ideal pest control method and that reliance on a single tactic can promote the development of resistant pest populations. Therefore, the goal is to identify the most effective control strategy by combining chemical, biological, genetic and physical techniques to regulate a specific pest12. The greenhouse whitefly, T. vaporariorum (Westwood, 1856) is considered one of the main pests associated with horticultural crops in protected environments in Uruguay, with tomato (Solanum lycopersicum L.) being one of the most affected 13)(14) . After several years of conventional chemical control, efforts have been made to reduce its populations through the use of the zoophytophagous predator Tupiocoris cucurbitaceus (Hemiptera: Miridae) 15)(16) 17, and the entomopathogenic fungus Beauveria bassiana (Balsamo) Vuillemin, with the aim of developing complementary, sustainable, and environmentally friendly control methods 18)(19) . Another major pest of tomato crop is the tomato leafminer Phthorimaea absoluta (Meyrick) (Lepidoptera: Gelechiidae), which causes damage to foliage, shoots flowers and fruits, both in open-field and greenhouse conditions20. Improving tomato production in the country and promoting the sustainability of production systems require the incorporation of new technologies that contribute to keep this pest population below economically damaging thresholds.
In this context, the objectives of this study were to evaluate the efficacy of T. cucurbitaceous and B. bassiana in controlling whitefly, to optimize their release strategy, and to evaluate their dispersal in the production system. For P. absoluta, the use of pheromone traps was evaluated to facilitate monitoring of the insect population in the crop, support decision-making, and assess the potential of mass trapping as a control strategy in tomato. This study presents the most recent research and advances in the biological control of two major tomato pests and the development of control tools compatible with natural enemies, naturally present or released into cropping systems and their environments in our country.
2. Materials and Methods
To achieve the stated objectives, tomato crops were established in experimental greenhouses located at the Entomology Unit, Faculty of Agronomy (Uruguay), where trials were conducted on the efficacy of the predator and the entomopathogenic fungus in controlling whitefly. Under commercial greenhouse conditions, T. cucurbitaceus was released in the horticultural region of Salto (Uruguay), where the method and timing of release, and its dispersion throughout the production system, were evaluated. In commercial tomato greenhouses in Canelones (Uruguay), the placement of traps with sex pheromone for controlling tomato moth was evaluated.
2.1 Effect of Tupiocoris cucurbitaceus on the Control of Trialeurodes vaporariorum in Experimental Greenhouse Tomato Crop
The study was conducted in a 50 m2 experimental greenhouse at the Agronomy Experimental Station in Salto -Uruguay (31°23'12" S 57°42'52" W), in the spring-summer 2017-2018 season. Six cages of 4 m2 each were set up and covered with mesh (MC Agryl P 17) to prevent insect movement. In each cage, ten Elpida variety tomato plants were placed in 5-lt plastic pots. These were planted at a density of 2.5 plants/m2 and managed according to standard tomato cultivation practices in the area. A localized automated irrigation system was installed for fertilization.
Two treatments were established: one with the release of T. cucurbitaceus and a control treatment without predator release. A completely randomized experimental design was used, with two treatments and three replicates. Each cage was a replicate of each of the treatments and contained 10 plants each21. Previously, the plants were infested with 25 whitefly adults (T. vaporariorum) per plant and, once they were established, the predators were released at a density of 0.6 Tupiocoris/m 2(22) . T. cucurbitaceus individuals used were sourced from Biobest (Argentina), in 125 ml flasks containing 500 adult individuals mixed with vermiculite. The release was carried out using an alternating plant schedule within the 10-plant plots, on leaves and flowers. The release was carried out 30 days after transplanting, once the establishment of whiteflies in the crop was confirmed. The temperature and relative humidity in the greenhouse were recorded using a HOBO sensor (H8 RH/Temp Loggers, Onset Computer Company, Bourne, MA, USA).
The variables evaluated were number of whitefly eggs, nymphs and adults per plant, as well as the number of T. cucurbitaceus adults per plant. Prior to the release of predators, the first whitefly sample (M0) was taken to record the population level in each plot. Two days after the introduction of the control agents, the second sample (M1) was taken, followed by weekly samples until the end of the experiment (M6). Sampling began in plots without T. cucurbitaceus to avoid mechanical transfer of predators between plots. In each sample, 10 leaves and 5 flowers were randomly removed from each plot. The leaves were placed in plastic bags, and the flowers were placed in flacks containing 70% alcohol for further processing in the laboratory. The number of whiteflies and predators, discriminating between mature and immature forms, was counted under a stereoscopic microscope (Nikon SMZ 1B- X 35) in the laboratory.
The variables eggs, nymphs, and adults were analyzed using repeated-measures ANOVAs for the six dates after the mirid's release. The log transformation (× +0.5) was used in all cases. The assumptions of homoscedasticity and normality were met.
2.2 Effect of Tupiocoris cucurbitaceus on the Control of Trialeurodes vaporariorum in Commercial Greenhouse Tomato Crops
Population monitoring of T. vaporariorum and its biocontrol agent T. cucurbitaceus was carried out in commercial greenhouse tomato crops in the horticultural region of Salto, Uruguay (31°23'12" S 57°42'52" W). In this context, two short-cycle tomato production periods were evaluated: one in autumn (February-June 2020) and another in spring (August to December 2019). The study was conducted in four commercial tomato greenhouses; each with a surface area of 2,381 m2 (113.4 × 21 m). The greenhouses followed the same sequence of agricultural practices for tomato cultivation, with differences in the dates of cultivation. T. cucurbitaceus individuals were released in two of these green-houses (the others were controls). From the crop transplant and once the presence of T. vaporariorum was confirmed, a weekly monitoring of their populations began and continued until the harvest.
For this, each greenhouse was subdivided into 20 quadrants and, in a plant of each quadrant, the number of adults of T. vaporariorum in the upper three leaves of the plant was counted weekly. Two weeks after transplanting and when the plants were at least 30 cm tall, mesh cages were set up with three tomato plants inside, at a rate of six cages per 1,000 m2 of greenhouse. Thirty T. cucurbitaceus individuals were released into each cage per plant, and eggs of Ephestia kuehniella Zeller (Lepidoptera, Pyralidae) were added to the leaves as a food supplement for predators. The egg dose was 1 g per 1,000 m2 of greenhouse during the first two weeks; the dose was doubled during the following two weeks, and tripled during the following two weeks. The cages remained closed for three weeks and then opened. To monitor predators, the number of T. cucurbitaceus was counted on the same leaves collected for T. vaporariorum monitoring.
For the analysis, ANOVA implemented in the R computing environment (R Development Core Team, 2014) was used. The normality of experimental errors as well as the homoscedasticity were checked. When the ANOVA reflected a significant difference, means were separated using the Tukey test.
2.3 Evaluation of the Dispersion of T. cucurbitaceus in the Greenhouses Where it was Introduced
To enable the commercial use of this predator, it is essential to evaluate whether its abundance increases in the surroundings of tomato greenhouses following its release, compared to conditions without release. For this purpose, studies were conducted to determine if T. cucurbitaceus disperses beyond the greenhouses where it was introduced and whether it persists throughout the seasons.
The predator is released into greenhouses using the inoculation-enhancement biological control method. Since this is a native species23 present in our production systems, it is not possible to attribute its presence and abundance to releases when data are collected far from the greenhouses and well after the release period. Therefore, the study was conducted from the time prior to release (day 0) until 30 days after, both inside and outside the greenhouses, with and without the release of the biocontrol agent.
The evaluations were carried out on two farms in the horticultural region of Salto, Uruguay (31°23'12" S 57°42'52" W), and covering four tomato greenhouses: two from the biological control program and two conventional pest control; each with a surface area of 2,381 m2 (113.4 × 21 m).
Two types of sampling were conducted: inside, which recorded the presence of the predator and whiteflies in the greenhouses, and outside, which recorded the presence of the mirid along transects extending in the four cardinal directions, at distances of 2 and 10 m from the greenhouse edge, 60 seconds per point with an entomological vacuum suction machine. For sampling inside, each greenhouse was subdivided into 20 quadrants and two leaves were randomly extracted from one plant per quadrant to determine the presence or absence of T. cucurbitaceus. Sampling was carried out in all greenhouses on day 0 (before release) and at 2, 5 and 30 days after the release of the predator. In the processing of the collected samples, a distinction was made between the different species of Miridae (Hemiptera) present in Uruguay24.
2.4 Evaluation of the Efficacy of Beauveria bassiana in the Control of Trialeurodes vaporariorum in an Experimental Tomato Greenhouse
The efficacy trial of B. bassiana was conducted in an experimental aluminum-frame greenhouse with a polyethylene roof, covering an area of 14 m2 and protected with 50-mesh anti-aphid netting. Inside the greenhouse, six voile cages with closures were installed to allow observation and simplify sample collection. Each cage contained six tomato plants of the Elpida variety. A completely randomized design was used, with three treatments and two repetitions per treatment. The control treatment (C) consisted of whitefly release followed by the application of distilled water. The first treatment with the entomopathogenic fungus (D1) consisted of the release of whiteflies and the subsequent application of B. bassiana at a dose of 200 g per 100 L of water. The second fungal treatment (D2) followed the same procedure, but with a higher concentration of B. bassiana at 300 g per 100 L of water. The fungal solution was prepared with the SUMA product (Registration No. 5439) as an adjuvant at a dose of 250cc/100L.
Applications of the entomopathogenic fungus were carried out weekly (11 weeks) following the label recommendations and using the doses indicated, always after 6:00 pm to avoid high temperatures. For the control treatment, applications with distilled water were made under the same conditions as the Beauveria treatments, using a different sprayer designated exclusively for that purpose. Applications began one week after the release of T. vaporariorum adults (30/11/23).
Weekly sampling was conducted to quantify the population of T. vaporariorum and the number of individuals infected by the fungus for 11 weeks. Monitoring was realized on tomato leaves, counting eggs, nymphs and adults of T. vaporariorum. For immature stages, in each cage three plants were randomly selected, and in each plant two leaflets were sampled, one of upper stratum and other from the lower stratum25. The tips of the same plants were evaluated, counting the number of adults on the three upper leaves that were fully unfolded.
Individual variables were analyzed using the Infostat program (with an R plugin)26, comparing the number of individuals (eggs, nymphs, adults, and total whitefly individuals) from the control and Beauveria treatments. Data were analyzed using generalized linear mixed models (MLGM) and fitted to the Poisson distribution. Means were compared using the Di Rienzo-Guzmán-Casanoves test (DGC ˂0.05). The percentage of control was calculated using Abbot's formula (Abbot, 1925) for the last three evaluation dates of the trial.
2.5 Monitoring and Mass Trapping of P. absoluta Using Pheromone Traps
In this study, to evaluate the effectiveness of mass traps against the tomato leaf miner, water traps equipped with Tuta Pro Caps® pheromone dispensers were installed (Figure 1)27. The attractant consists of a cylindrical syringe containing 0.5 mL of sex pheromone. For mass trapping, the manufacturer recommends placing 40 traps per hectare under greenhouse conditions, spaced 15 meters to ensure a 7-meter radius of action per trap and avoid interference between attractants.
The experiment was carried out in two tomato (Lycopersicon esculentum L.) greenhouses of the Eterei variety transplanted on 9/9/22, located in the town of Villa García (34°46' 54.96" S 56° 4' 52.99" W). The start date of field activities was 26/9/22 with the installation of the traps and the evaluations culminated on 26/1/23.

Figure 1: Greenhouse with mass trapping using sex pheromones (left) and control greenhouse without mass trapping (right)
The traps consist of cylindrical containers containing a capture liquid composed of water and a few drops of soap. This liquid breaks the surface tension and allows the males to submerge themselves when landing28. They were placed 30-40 cm above the ground in plastic boxes, at a density of 30 traps/ha. A weekly monitoring of P. absoluta was carried out, collecting males captured in the water traps placed inside the greenhouse where mass trapping was implemented. Additionally, weekly assessments were conducted to record the number of plants exhibiting galleries (incidence) and the number of galleries per plant (severity), both in the greenhouse with mass trapping and in the control greenhouse. For in-plant monitoring, the methodology was the one proposed by Polack and Mitidieri29. Until the plant reaches 1 meter in height, all leaflets are sampled; after exceeding 1 meter, the top 10 leaves are monitored until the plant reaches 1.70 m in height. From this point on, the 10 leaves in the middle stratum of the plant are monitored. Both trials (treatment and control) started 20 days after the crop was transplanted and P. absoluta was recorded in the greenhouses.
The number of plants with live galleries as an incidence indicator was analyzed using the Kruskal Wallis test. The cumulative number of live galleries per plant, as a severity pest damage indicator, was analyzed using the Kruskal Wallis test globally and by monitoring date. Both indicators were analyzed using the Infostat software of Di Rienzi and others26.
3. Results
3.1 Effect of T. cucurbitaceus on the Control of the Whitefly T. vaporariorum in Tomato Crop under Experimental Greenhouse Conditions
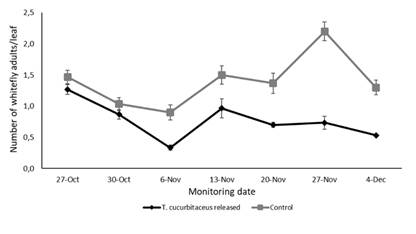
Monitoring of T. vaporariorum populations (eggs, nymphs and adults) on leaves showed significant variation over the sampling period (F1,60= 54.208, p < 0.001), with a marked decline in the plots where predator releases were conducted (Figure 2, Figure 3 and Figure 4). The number of eggs (F1,20 = 28.201, p = 0.006044), nymphs (F1,20 = 101.04, p = 0.000551) and adults (F1,20 = 49.245, p = 0.002172) were significantly lower in the plots with predator presence compared to the control plots, where T. cucurbitaceus was never detected. By the end of the experiment, the plots where the mirid was released exhibited a reduction of approximately 80% in whitefly population levels compared to the control treatment. No additional predator releases or alternative control methods were necessary once T. cucurbitaceus established in the crop.
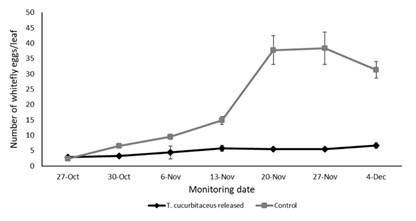
Figure 2: Number of whitefly eggs on tomato leaves in plots with release of T. cucurbitaceus and without release of T. cucurbitaceus (control) from October to December 2023
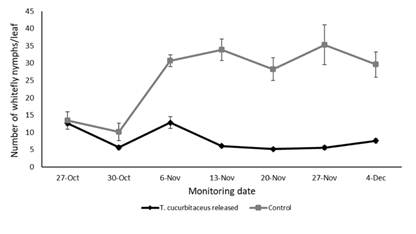
Figure 3: Number of whitefly nymphs on tomato leaves in plots with release of T. cucurbitaceus and without release of T. cucurbitaceus (control) from October to December 2023
3.2 Assessment of T. cucurbitaceus for the Control of T. vaporariorum in Commercial Tomato Greenhouses
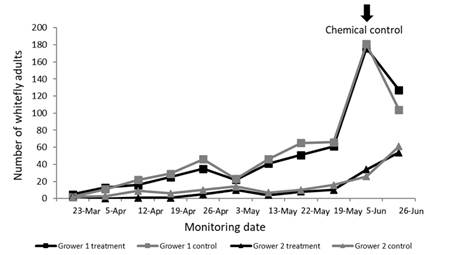
During the final weeks of the crop cycle (6/11-18/11/19), pest population levels were considerably higher in the control greenhouse compared with the greenhouse where the predator was released, in the case of Grower 1 (p < 0.001) (Figure 5). In contrast, no significant differences in whitefly populations were observed between treatments for Grower 2 (p = 0.472) (Figure 5). Notably, the predator demonstrated greater efficacy during the short spring cycle (Figure 5) than during the autumn cycle (p < 0.001) (Figure 6). In addition, in the autumn cycle, no differences were observed between control and treated greenhouses for either of the two growers (p = 0.087) (Figure 6).
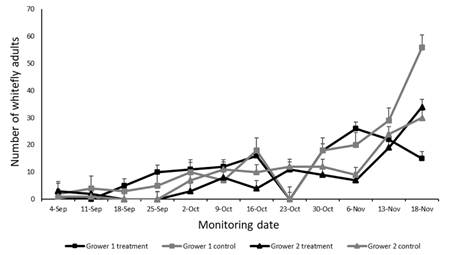
Figure 5: Number of adult whiteflies by sampling date and grower, with (treatment) and without release of Tupiocoris cucurbitaceus (control), in tomato greenhouses during a short spring cycle (from September to November 2019)
3.3 Evaluation of the Dispersion of T. cucurbitaceus in the Greenhouses where it was Introduced
The population levels of the mirid in Grower 1 showed an increasing trend inside the greenhouses until the end of May. Subsequently, a decline in population was observed, possibly due to the application of chemical insecticides aimed at controlling other crop pests (Figure 7). The absence of the predator in Grower 2 is due to the low whitefly population levels that remained constant throughout the crop cycle (Figure 7).
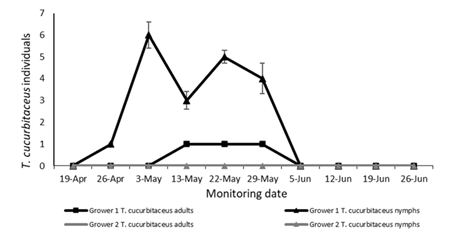
Figure 7: Number of T. cucurbitaceus nymphs and adults by sampling dates and grower in greenhouse tomato crops in Salto (from April to June 2019)
No individuals of T. cucurbitaceus were detected outside the greenhouses at the distances and time established in the study. After the experiment concluded, sampling was conducted on the farms of both producers from July to September. During this time, T. cucurbitaceus individuals were found in red pepper greenhouses located approximately 200 meters from the tomato crops where the release took place. However, due to the time elapsed and the fact that the mirid has been observed naturally in areas where releases have never been carried out, it cannot be confirmed that these individuals migrated from the release site.
3.4 Evaluation of the Efficacy of the Entomopathogenic Fungus B. bassiana in the Control of T. vaporariorum in an Experimental Greenhouse Tomato Crop
The first whitefly eggs were observed in the control treatment, whereas oviposition in the plots treated with B. bassiana was only recorded from the third week after the appearance of the control (Figure 8). The number of eggs per leaf was significantly higher in the control treatment than in the plots treated with the entomopathogen (F = 3.47, p = 0.0322).
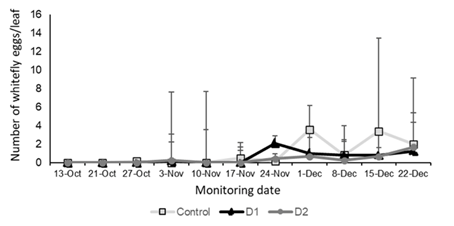
Figure 8: Number of T. vaporariorum eggs per sampled leaf area (individuals/leaf in 4.9 cm2) in the treatments with B. bassiana application (D1 and D2) and the control treated with distilled water
Nymph levels per leaf increased steadily until the end of the experiment in the control treatment, reaching 9 nymphs per leaf. In contrast, in the treatments with fungal application, nymph levels remained below 3 nymphs per leaf (Figure 9). The control showed statistically higher values than the treated plots (F = 8.15, p = 0.0003).
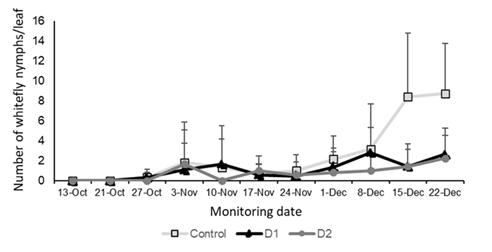
Figure 9: Population levels of T. vaporariorum nymphs per sampled leaf area (individuals/leaf in 4.9 cm2), in the treatments with B. bassiana application and the control treated with distilled water
The number of whitefly adults observed on leaflets remained below 0.5 adults per leaf in all treatments, except for the control, where a population increase was recorded starting from December, reaching an average of 3.25 adults per leaf (Figure 10). Adult counts per leaf also showed significant differences among treatments, with high number in the control treatment (F = 12.20, p < 0.0001).
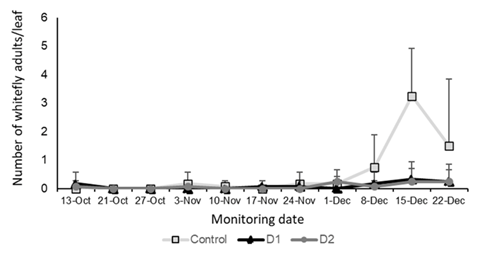
Figure 10: Population levels of T. vaporariorum adults per sampled leaf area (individuals/leaf in 4.9 cm2), in the treatments with B. bassiana application and the control treated with distilled water
Considering the total number of whitefly individuals (eggs, nymphs and adults), an increase was observed in the control treatment from week 7, reaching a maximum average of 15.08 individuals per leaf at week 9. On the contrary, in the plots where the fungus was applied (D1 and D2), the maximum average recorded was 4 individuals per leaf at week 8. The control treatment showed significantly higher whitefly populations than the treatments where B. bassiana was applied (F = 10.27, p < 0.0001) (Figure 11).
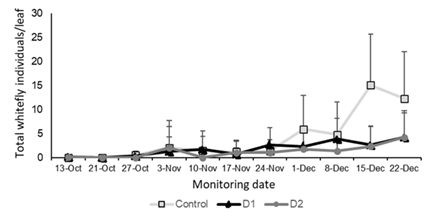
In the monitoring of adults on the plant, a sustained increase in the number of individuals was observed in the control, reaching a maximum record of 5.67 individuals per plant. In contrast, in the treatments involving fungal application, the levels never exceeded 1 individual per plant (Figure 12). Monitoring of adults on the top three leaves of the plant also revealed significant differences between the control and the treatments with B. bassiana application (F = 13.78, p < 0.0001).
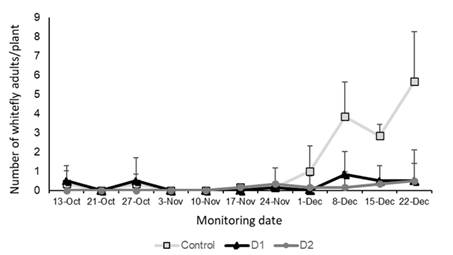
Figure 12: Adult population levels of T. vaporariorum per plant in treatments with B. bassiana application and the control with distilled water application
The best control levels during the period in which this study was conducted were achieved with dose 1 (D1: 200 g/100 L), as a higher dose did not result in greater control under these conditions. In fact, in some cases, the highest dose yielded lower levels of control. The percentage of control achieved at the foliage sampling level, calculated from the total number of individuals, was greater than 65%, while the percentage of control achieved when monitoring the three upper leaves of the plant was greater than 91% (D1) and 85% (D2). Therefore, this strain appears to be a promising tool for controlling this pest.
The entomopathogenic fungus Beauveria bassiana demonstrated strong efficacy against T. vaporariorum, with both doses tested significantly reducing pest populations. No differences were observed between doses, suggesting that lower concentrations may be sufficient under controlled conditions.
3.5 Monitoring and Mass Trapping of P. absoluta Using Pheromone Traps
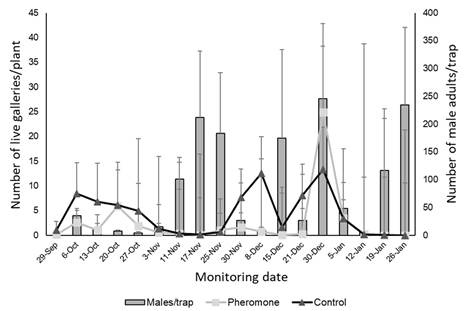
Captures occurred from the time the traps were placed until the end of the experiment, indicating that the traps remained functional throughout the evaluation period (Figure 13).
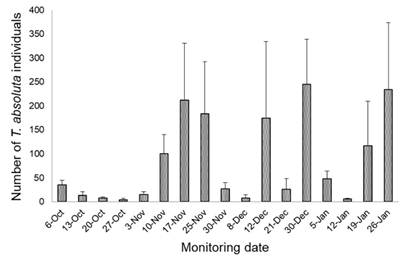
Figure 13: Average of P. absoluta males captured in four pheromone traps deployed inside the greenhouse with mass trapping (from October 2022 to January 2023)
Population fluctuations in the number of males captured in traps revealed several peaks that could correspond to different generations of the pest. The first peak was recorded on November 10, while subsequent peaks were more frequent and could reflect shorter generational cycles, possibly due to rising temperatures or overlapping generations (Figure 13).
Monitoring conducted directly on plants indicated a higher incidence of the pest in the control greenhouse (without mass trapping) compared with the treatment greenhouse with mean values of 69% and 40%, respectively (H = 5.57, p = 0.0173) (Figure 14). Notably, on December 30, an incidence of 100% of plants with galleries was recorded in both the control and the treated greenhouses. This high incidence in the treated greenhouse coincided with a saturation of the trap on that date, with an average capture rate of 245 males per trap.
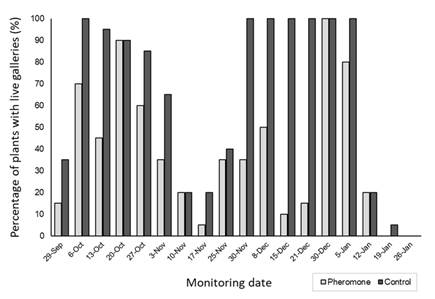
Figure 14: Incidence of plants with live galleries in the greenhouse with mass trapping (grey bars) and in the greenhouse without trapping (black bars) (from September 2022 to January 2023)
Severity followed a similar pattern, with a higher number of live galleries per plant recorded in the control greenhouse compared with the greenhouse with mass trapping (Figure 15 and Figure 16) (H = 59.21, p < 0.0001). However, on December 30, this trend was reversed, with a great number of galleries per plant observed in the treated greenhouse (H = 4.34, p = 0.0370). As mentioned before, this date coincides with a time of saturation of the traps.
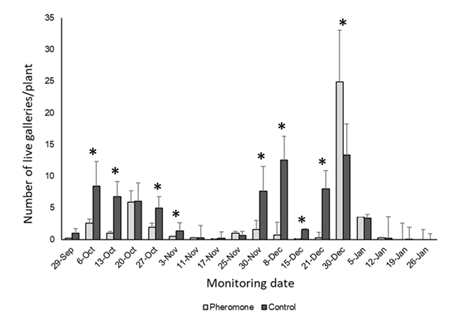
Figure 15: Severity expressed as live galleries per plant in greenhouse with mass trapping (grey bars) and control greenhouse (black bars)
4. Discussion
The release of natural enemies in horticultural greenhouses reduce insecticide use, keeping pest population levels below damage thresholds and production within expected standards. In this context, the development of this initiative has opened new avenues for research on the use of entomophagous insects in Uruguay. It is worth noting that healthier, pesticide-free production improves quality of life, protects the health of growers and enhances environmental sustainability for the entire community, including end consumers.
Results under greenhouse experimental conditions demonstrated the high efficacy of the mirid T. cucurbitaceus in controlling T. vaporariorum populations in tomato. This predator is highly polyphagous and well adapted to the environmental conditions typical of the horticultural regions of Uruguay. It also contributes to the control of other common crop pests, including eggs and larvae of the tomato leafminer, thrips, aphids, and spider mites30. Therefore, with proper management, this agent can be effectively integrated into an IPM for tomato crops.
However, as zoophytophagous insects, they can cause damage to plant tissue in the absence of prey31. Recent studies have evaluated the phytophagous capacity of T. cucurbitaceus in tomato plants, demonstrating that this species does not cause significant damage or yield losses 15)(16) . However, although T. cucurbitaceus is a very effective predator for controlling whitefly, its establishment is slow and requires the population to reach high levels before the onset of pest infestation to ensure effective control. Furthermore, its effectiveness in pest control is closely related to the success of establishment in the crop; when establishment is null or insufficient, it is often a major cause of failureMitidieri29. Nevertheless, increasing the release doses makes its use economically feasible only in high-yielding production systems.
Another important aspect to keep in mind is its difficulty to remain inside the greenhouse when tomato plants are still small and the pest population is low. This is due to its behavior, which is highly dependent on the predator/prey ratio. In semi-open greenhouse conditions, surrounded by other crops that harbor prey species, and given that it is a polyphagous predator32, individuals tend to disperse in search of alternative prey. Although T. cucurbitaceus occurs naturally in our production systems, high densities of the mirid must be quickly achieved in tomato crops to prevent whitefly outbreaks. Therefore, the method and timing of release must be adjusted in commercial greenhouses. In the research trials conducted, the predator successfully established itself in the tomato crop, although its establishment required several weeks to reach the desired population levels. Given its slow establishment and the short crop cycle, by the time the natural enemy establishes itself and begins to control the pest, there are only a few weeks left before harvest. Based on these results, we believe its use is best suited for long-cycle tomato crops. Furthermore, it would be crucial to evaluate the interaction of T. cucurbitaceus with other biocontrol agents present in the system, such as parasitoids (Encarsia sp., Eretmocerus sp.) or entomopathogens (B. bassiana), in order to develop an integrated control strategy for T. vaporariorum. Additionally, the B. bassiana strain evaluated proved to be effective in controlling this whitefly under experimental conditions. Additional studies in commercial greenhouses are required to adjust their application.
Effective control of P. absoluta was observed using mass trapping at low pest densities. At high population levels, trap saturation occurs, as occurred in our study, which reduces the effectiveness of mass trapping by failing to prevent mating. These results highlight the importance of integrating chemical treatments under these conditions, a decision that can be guided by trap capture dynamics. However, it is worth noting that despite high capture rates, the severity of damage in the pheromone-treated greenhouse remained lower than in the control. Monitoring male captures combined with evaluation of plant galleries is a key tool for determining the need for additional interventions, particularly when trap saturation occurs. In fields with a history of high P. absoluta infestations, mass trapping should be combined with preventive strategies, such as treatments based on beneficial microorganisms or entomophagous agents like T. cucurbitaceus, to prevent the pest's population growth. If these methods are insufficient, curative treatments with low-impact products should be considered.
Considering that T. cucurbitaceus is capable of preying on the eggs and larvae of the tomato leafminer P. absoluta29, its combined use with mass trapping could offer effective pest control and represents an additional aspect worth evaluating. When implementing biological control, it is necessary to consider its compatibility with other integrated pest management (IPM) strategies, as these may indirectly affect the natural enemies targeted for conservation and promotion. National-level data are needed to plan and monitor IPM programs that incorporate low environmental impact tools. Additionally, the development of public policies that support growers in the implementation of these long-term control programs is crucial, as is the involvement of private companies in the commercialization of biological products to promote the development of more sustainable horticulture.
5. Conclusions
This study highlights the potential of integrating multiple environmentally friendly strategies for the effective control of key tomato pests in greenhouse conditions.
The mirid Tupiocoris cucurbitaceus proved to be a highly effective predator of Trialeurodes vaporariorum. However, its successful establishment depends on adequate release strategies, environmental conditions, and pest presence, making it more suitable for long-cycle crops. Further research on food supply and banker plants systems should be developed in order to explore different potential ways to promote T. cucurbitaceus establishment after release.
The entomopathogenic fungus Beauveria bassiana demonstrated strong efficacy against T. vaporariorum, with both doses tested, significantly reducing pest populations. Further evaluations under commercial settings are needed to optimize its application and assess its compatibility with zoophytophagous predators.
Mass trapping with Tuta Pro Caps® pheromone was effective in reducing P. absoluta populations, particularly at low infestation levels. However, under high pest pressure, trap saturation limited its efficacy, highlighting the need for complementary control measures such as insecticide applications or biological agents, particularly in areas with a history of high pest incidence. A regional approach to managing this pest through mass trapping with pheromones would be the most recommended.

